
5. Морфология бактерий. Отличия прокариотов от эукариотов. Основные формы бактерий.
Пo форме клеток м/о собственно подразделяются на:
A) Шаровидные (кокки) правильной формы сферической или овальной формы:
а) микрококки располагаются беспорядочно.
б) диплококки — располагаются парами
в) стрептококки цепочки в виде пакетов из четырех кокков
г) тетракокки — располагаются в виде пакетов из четырёх кокков
д) сарцины — располагаются в виде пакетов из восьми кокков.
е) стафилококки — скопление кокков в форме виноградных гроздей.
Патогенные бактерии чаще всею представлены стафилококками, стрептококками, реже микрококками. Спор и жгутиков патогенные кокки не образуют. Капсулы имеют лишь S.pneumoniae. Стафилококки и стрептококки — грамположительные, диплококки — грамотрицательные.
Б) Палочковидные бактерии цилиндрической формы, различаются по расположению, размерам, форме клеток и их концов.
а) в виде одиночных клеток
б) диплобактерии — располагаются парами
в) стрептобактерии — располагаются в виде цепочек
Патогенные палочковидные бактерии располагаются в основном беспорядочно, попарно, многие палочковидные бактерии имеют жгутики и капсулы, бациллы и клостридии имеют споры. Коккобактерии и палочки окрашиваются грамотрицатсльно, бациллы, клостридии, корине- и микобактерии -грамположительно.
B) Извитые формы бактерий — представлены изогнутыми палочками с
а) одним изгибом (холерный вибрион)
б) с несколькими изгибами (кампилобактерии)
Спор и капсул вибрионы не образуют. В мазке напоминают паутинку, окрашиваются грамотрицательно. Размеры бактерий измеряются в мкм, их органелл — в нм.
Отличия прокариот от эукариот
1. У прокариот отсутствуют мембраны, ограничивающие органеллы бактериальной клетки(ядро. митохондрии, рибосомы) от цитоплазмы. Из мембран имеется только цитоплазматическая мембрана.
2. Ядро прокариот (нуклеоид) фибриальной структуры, ядерная оболочка отсутствует.
3. У прокариот отсутствуют митохондрии, хлоропласты, КГ. ЭПС.
4. Окислительно-восстановительные фрагменты локализованы в мезосомах (производных цитоплазматической мембраны)
5. У прокариот отсутствует митоз, размножаются путем бинарного деления.
6. Прокариоты имеют гаплоидный геном.
7. Отсутствует клеточный центр
8. Внутриклеточные перемещения цитоплазмы и амебоидное движение для прокариот нетипичны.
Специфические черты М/О
1. Малые размеры, масса, объем и относительная простота строения.
2. Чрезвычайно высокие темпы размножения
3. Большое разнообразие способов получения энергии и путём обмена веществ, широкий спектр конечных продуктов метаболизма.
4. Способность к биодеструкции практически всех естественных и искусственных веществ.
5. Чрезвычайно высокая степень адоптации как результат высоких темпов изменчивости.
6. Массовая популяция и повсеместное распространение.
6. Структура и функции поверхностных образований бактериальной клетки. Капсула. Методы выявления.
Бактериальная клетка окружена внешней оболочкой (рис. 3.2), которая состоит из капсулы, капсулоподобной оболочки и клеточной стенки. От их состава зависит способность клетки воспринимать анилиновые красители (тинкториальные свойства). Капсулы в зависимости от степени выраженности подразделяют на микро- и макрокапсулы. Первые обнаруживаются только при электронно-микроскопическом исследовании в виде микрофибрилл из мукополисаха-ридов, которые тесно прилегают к клеточной стенке. Макрокапсулы представляют собой выраженный слизистый слой, снаружи покрыва ющий клеточную стенку. Он состоит из полисахаридов и редко из полипептидов (например, у сибиреязвенных бактерий). Как правило макрокапсулу образуют немногие виды патогенных бактерий (пнев мококки и др.) при неблагоприятных условиях среды, например в организме животных или человека. Однако у некоторых видов (клебсиеллы пневмонии) макрокапсула обнаруживается постоянно.
Капсулоподобная оболочка — липидо-полисахаридное образование, сравнительно непрочно связанное с поверхностью клетки, вследствие чего в отличие от капсулы может выделяться в окружающую среду.
Капсула или капсулоподобная оболочка может быть покрыта экзополисахаридами, которые образуются из углеводов окружающей среды под действием бактериальных ферментов. При этом глюканы и леваны обеспечивают прилипание бактерий к разным поверхностям, часто гладким.
Капсула несет различные функции:
1. Защитная, предохраняя клетку от неблагоприятных условий среды обитания,
2. адгезивная, способствуя «прилипанию» к поверхносш (рецепюрам) клетки хозяина.
3. Часто патогенные и антигенные свойства. Непатогенные бактерии также могут образовывать макрокапсулу, выполняющую, по-видимому, только защитную функцию.
studfiles.net
4. Морфология, ультраструктура и химический состав бактерий.
Морфологические свойства бактерий. Бактерии — микроорганизмы, не имеющие оформленного ядра (прокариоты).
Для бактерий характерны четыре основные формы: сферическая (шаровидная), цилиндрическая (палочковидная), извитая и нитевидная.
Размеры бактерий колеблются от 0,1 до 10 мкм.
Капсула – необязательный полисахаридный или полипептидный компонент, выполняющий:
1. Обусловливает форму клетки. 2. Защищает клетку от механических повреждений извне и выдерживает значительное внутреннее давление. 3. Обладает свойством полупроницаемости, поэтому через нее избирательно проникают из среды питательные вещества. 4. Несет на своей поверхности рецепторы для бактериофагов и различных химических веществ.
Жгутики – необязательный белковый компонент (флагеллин), функция – движение, антиген.
Микроворсинки или пили – необязательный белковый компонент, функция: пили адгезии, конъюгативные пили.
Клеточная стенка – обязательный белковый компонент, выполняющий функции: защитную, барьерную, транспортную, рецепторную, выделительную, антигенную, деление, спорообразующую.
Гр+ — 40 слоев, + тейхоевые кислоты, + РНК, Mg.
Гр- — имеют внешнюю мембрану, 2 слоя, ЛПС слой.
Мезосомы – аналоги МТХ, участвуют в делении клетки.
Рибосомы – синтез белка.
Нуклеоид – кольцевая молекула ДНК, несущая генетическую информацию.
Споры – образуют только бациллы и клостридии. Маловодны, много Са, много жирных кислот, окраска по Цилю-Нильсену.
Для бактерий рекомендованы следующие таксономические категории: класс, отдел, порядок, семейство, род, вид. Название вида соответствует бинарной номенклатуре, т. е. состоит из двух слов.
L-формы — это бактерии, полностью или частично лишенные клеточной стенки, поэтому имеют своеобразную морфологию в виде крупных и мелких сферических клеток. Способны к размножению.
Цитоплазматическая мембрана располагается под клеточной стенкой (между ними — периплазматическое пространство). По строению является сложным липидобелковым комплексом.
Функции цитоплазматической мембраны: 1. Является основным осмотическим и онкотическим барьером. 2. Участвует в энергетическом метаболизме и в активном транспорте питательных веществ в клетку, так как является местом локализации пермеаз и ферментов окислительного фосфорилирования. 3. Участвует в процессах дыхания и деления. 4. Участвует в синтезе компонентов клеточной клетки (пептидогликана). 5. Участвует в выделении из клетки токсинов и ферментов.
5. Основные отличия прокариот и эукариот, прокариот и вирусов.
Прокариоты отличаются от эукариот по ряду основных признаков.
1.Отсутствие истинного ядра ядерной мембраны, гистонов. Хромосома в кольце,
2.Отсутствие развитой эндоплазматической сети, аппарата Гольджи.
3.Отсутствие митохондрий, хлоропластов, лизосом.
4.клеточная стенка – пептидогликан
5.Размножение бинарное в отличие от митоза и мейоза.
6. Значительно меньшие размеры
7. Тип дыхания как аэро так и анаэробное
Вирусы — это всегда внутриклеточные паразиты. Они способны размножаться только в чужой клетке, потому что сами состоят только из одного типа нуклеиновой киcлоты и белковой или белково-липидной оболочки.
Вирусы собственного обмена веществ не имеют. Они внедряются в клетку и заставляют ее изготавливать копии вируса. Идет производство копий нуклеиновой кислоты и копий вирусных белков, из которых в конце собирается новая вирусная частица. При этом клетка чаще всего гибнет из-за прекращения продукции собственных белков, накопления токсических вирусных компонентов и повреждения клеточных лизосом
studfiles.net
5. Морфология бактерий. Отличия прокариотов от эукариотов. Основные формы бактерий.
Пo форме клеток м/о собственно подразделяются на:
A) Шаровидные (кокки) правильной формы сферической или овальной формы:
а) микрококки располагаются беспорядочно.
б) диплококки — располагаются парами
в) стрептококки цепочки в виде пакетов из четырех кокков
г) тетракокки — располагаются в виде пакетов из четырёх кокков
д) сарцины — располагаются в виде пакетов из восьми кокков.
е) стафилококки — скопление кокков в форме виноградных гроздей.
Патогенные бактерии чаще всею представлены стафилококками, стрептококками, реже микрококками. Спор и жгутиков патогенные кокки не образуют. Капсулы имеют лишь S.pneumoniae. Стафилококки и стрептококки — грамположительные, диплококки — грамотрицательные.
Б) Палочковидные бактерии цилиндрической формы, различаются по расположению, размерам, форме клеток и их концов.
а) в виде одиночных клеток
б) диплобактерии — располагаются парами
в) стрептобактерии — располагаются в виде цепочек
Патогенные палочковидные бактерии располагаются в основном беспорядочно, попарно, многие палочковидные бактерии имеют жгутики и капсулы, бациллы и клостридии имеют споры. Коккобактерии и палочки окрашиваются грамотрицатсльно, бациллы, клостридии, корине- и микобактерии -грамположительно.
B) Извитые формы бактерий — представлены изогнутыми палочками с
а) одним изгибом (холерный вибрион)
б) с несколькими изгибами (кампилобактерии)
Спор и капсул вибрионы не образуют. В мазке напоминают паутинку, окрашиваются грамотрицательно. Размеры бактерий измеряются в мкм, их органелл — в нм.
Отличия прокариот от эукариот
1. У прокариот отсутствуют мембраны, ограничивающие органеллы бактериальной клетки(ядро. митохондрии, рибосомы) от цитоплазмы. Из мембран имеется только цитоплазматическая мембрана.
2. Ядро прокариот (нуклеоид) фибриальной структуры, ядерная оболочка отсутствует.
3. У прокариот отсутствуют митохондрии, хлоропласты, КГ. ЭПС.
4. Окислительно-восстановительные фрагменты локализованы в мезосомах (производных цитоплазматической мембраны)
5. У прокариот отсутствует митоз, размножаются путем бинарного деления.
6. Прокариоты имеют гаплоидный геном.
7. Отсутствует клеточный центр
8. Внутриклеточные перемещения цитоплазмы и амебоидное движение для прокариот нетипичны.
Специфические черты М/О
1. Малые размеры, масса, объем и относительная простота строения.
2. Чрезвычайно высокие темпы размножения
3. Большое разнообразие способов получения энергии и путём обмена веществ, широкий спектр конечных продуктов метаболизма.
4. Способность к биодеструкции практически всех естественных и искусственных веществ.
5. Чрезвычайно высокая степень адоптации как результат высоких темпов изменчивости.
6. Массовая популяция и повсеместное распространение.
6. Структура и функции поверхностных образований бактериальной клетки. Капсула. Методы выявления.
Бактериальная клетка окружена внешней оболочкой (рис. 3.2), которая состоит из капсулы, капсулоподобной оболочки и клеточной стенки. От их состава зависит способность клетки воспринимать анилиновые красители (тинкториальные свойства). Капсулы в зависимости от степени выраженности подразделяют на микро- и макрокапсулы. Первые обнаруживаются только при электронно-микроскопическом исследовании в виде микрофибрилл из мукополисаха-ридов, которые тесно прилегают к клеточной стенке. Макрокапсулы представляют собой выраженный слизистый слой, снаружи покрыва ющий клеточную стенку. Он состоит из полисахаридов и редко из полипептидов (например, у сибиреязвенных бактерий). Как правило макрокапсулу образуют немногие виды патогенных бактерий (пнев мококки и др.) при неблагоприятных условиях среды, например в организме животных или человека. Однако у некоторых видов (клебсиеллы пневмонии) макрокапсула обнаруживается постоянно.
Капсулоподобная оболочка — липидо-полисахаридное образование, сравнительно непрочно связанное с поверхностью клетки, вследствие чего в отличие от капсулы может выделяться в окружающую среду.
Капсула или капсулоподобная оболочка может быть покрыта экзополисахаридами, которые образуются из углеводов окружающей среды под действием бактериальных ферментов. При этом глюканы и леваны обеспечивают прилипание бактерий к разным поверхностям, часто гладким.
Капсула несет различные функции:
1. Защитная, предохраняя клетку от неблагоприятных условий среды обитания,
2. адгезивная, способствуя «прилипанию» к поверхносш (рецепюрам) клетки хозяина.
3. Часто патогенные и антигенные свойства. Непатогенные бактерии также могут образовывать макрокапсулу, выполняющую, по-видимому, только защитную функцию.
studfiles.net
Как уже упоминалось, весь органический мир делится на две части; прокариоты и эукариоты. Рассмотрим их более побробно. Прокариоты не имеют ядра с мембраной, и генетический материал сосредоточен в нуклиотиде. Дезоксирибонуклеиновая кислота (ДНК) образует единственную нить, замкнутую в кольцо(генофор). Половой процесс отсутсвует, а обмен генетическим материалом осуществляется во время других процессов, называемых парасексуальными. Эукариоты — организмы с настоящим ядром, окруженным ядерной мембраной. Генетический материал заключен в хромосомах, состоящих из нитей ДНК и белков. Для эукариотов характерен типичный половой процесс с чередующимся слиянием ядер и редукционным делением; иногда размножаются без оплодотворения, но при наличии половых органов (партеногенез). Клетка имеет центриоли, митотическое веретено, пластиды, митохондрии и хорошо развитую эндоплазматическую мембранную систему. Деление клеток митотическое. Если имеются жгутики или реснички, то они очень сложно устроены. Атмосферный азот они не фиксируют, как правило, аэробы, редко вторичные анаэробы. Питание абсорбтивное или автотрофное, когда пища заглатывается и переваривается внутри организма. Имеются пищевые вакуоли. В лаборатории для определения вида микроорганизмов устанавливают его основные свойства: морфологию, рост, на питательных средах, биохимические свойства, патогенность и другое. По полученным данным проводят идентификацию по определению находят место микроба в классификационной таблице. Вирусы. Относятся они к облигатным внутриклеточным паразитам, их размеры варьируют в широких пределах – от 10 до 200 нм и более. Величину их определяют фильтрованием через фильтры с известным размером пор, центрифугированием в скоростных центрифугах и исследованием под электронным микроскопом. Риккетсии. Риккетсии относят к болезнетворным организмам, занимающим промежуточное положение между вирусами и бактериями. Они значительно меньше бактерий, но крупнее вирусов и едва заметны в световой микроскоп. Это, как правило, облигатные внутриклеточные паразиты размером от 300 до 2000 нм. По своему строению риккетсии сходны с бактериями и подразделяются на коковидные, полочковидные и нитевидные. Последние могут достигать длины 40 мкм.По строению напоминают грамотрицательные бактерии. Бактериофаги. Это вирусы, развивающиеся в микроорганизмах. Подобного рода вирусы распространены в природе повсюду, где встречаются бактерии. Микоплазмы (спироплазмы). Размеры микоплазм колеблются от 100 до 700 нм, спор они не образуют. Растут на сложных питательных средах с высоким осмотическим давлением. Колонии врастают в плотные среды. Отсутствие истинной клеточной оболочки (её заменяет 3-х слойная мембрана из стериновых лепидов) у микоплазм ведёт к выраженному полиморфизму — шаровидным, зернистым, колцевидным и нитевидным формам. Способность проникать через бактериальные фильтры, свидетельствует об их морфологической пластичности. Микоплазмы широко распространены в природе и имеют важное значение в патологии животных, птиц и насекомых, к которым относятся пчёлы. |
www.obnogka.ru
Признак | Прокариоты | Эукариоты | |
Особенности строения клеток | |||
Наличие ядра | Обособленного ядра нет | Обособленное ядро, отделенное от цитоплазмы двойной мембраной. | |
Число хромосом и их строение | У бактерий – одна кольцевая хромосома, прикрепленная к мезосоме, двуцепочечная ДНК, не связанная с белками гистонами. У цианобактерий – несколько хромосом в центре цитоплазмы. | Определенное число хромосом для каждого вида. Хромосомы линейные, двуцепочечная ДНК, связанная с белками гистонами. | |
Плазмиды | Имеются. | Имеются у митохондрий и у пластид. | |
Ядрышки | Отсутствуют. | Имеются. | |
Организация генома | До 1,5 тысяч генов. Большинство генов в единственной копии. | В зависимости от вида от 5 до 200 тысяч генов. У человека около 100 тысяч генов. Доля генов, представленных в нескольких копиях достигает 45 %, число копия одного гена может достигать нескольких тысяч. | |
Рибосомы | Мельче, чем у эукариот – от 70 S. Распределены в цитоплазме. Обычно свободные, редко связанные с мембранными структурами. Составляют до 40 % массы клетки. | Крупные, от 80 S. Находятся в цитоплазме в свободном состоянии или связаны с мембранами ЭПР. В пластидах и митохондриях содержатся рибосомы 70 S. | |
Одномембранные замкнутые органеллы | Отсутствуют. Их функции выполняют выросты клеточной мембраны. | Многочисленны: ЭПР, АГ, вакуоли, лизосомы. | |
Двухмембранные органеллы | Отсутствуют. | Митохондрии у всех эукариот, пластиды – у растений. | |
Клеточный центр | Отсутствует. | Имеется в клетках животных, грибов, растений (мхи и водоросли). | |
Мезосомы | Имеются у бактерий. | Отсутствуют. | |
Клеточная стенка | У бактерий содержит муреин, у цианобактерий – целлюлозу, пектиновые вещества, немного муреина | У растений – целлюлоза, у грибов – хитин, у животных клеточных стенок нет. | |
Признак | Прокариоты | Эукариоты | |
Капсула и слизистый слой | Имеются у некоторых бактерий. | Отсутствуют. | |
Жгутики | Простое строение, не содержат микротрубочек. Диаметр 20 нм. | Сложного строения, содержат микротрубочки. Диаметр 200 нм. | |
Размер клеток | Диаметр 0,5 – 5 мкм. | Диаметр до 50 мкм. Объем может превышать объем прокариотической клетки в тысячу и более раз. | |
Особенности жизнедеятельности клеток | |||
Движение цитоплазмы | Отсутствует. | Наблюдается часто. | |
Аэробное дыхание | У бактерий – в мезосомах, у цианобактерий – на ЦПМ. | В митохондриях. | |
Фотосинтез | Хлоропластов нет. У фотосинтезирующих бактерий и цианобактерий происходит на мембранах. | В хлоропластах у растений. | |
Фагоцитоз и пиноцитоз | Отсутствует из-за наличия жесткой клеточной стенки. | Только у животных. | |
Спорообразование | Некоторые бактерии обладают способностью образовывать спору, предназначенную для перенесения неблагоприятных условий среды. | Свойственно растениям и грибам, предназначены для размножения. | |
Способы деления клетки | Равновеликое бинарное деление, редко почкование. Митоз и мейоз отсутствуют. | Митоз, мейоз, амитоз. | |
studfiles.net
2 Прокариотические организмы. Бактерии.
2 Прокариотические организмы. Бактерии.
Бактерии — это мельчайшие прокариотические организмы, имеющие клеточное строение. По причине микроскопических размеров клеток от 0,1 до 1030 мкм бактерии получили название микробов или микроорганизмов.
Бактерии живут в почве, воде, воздухе, снегах полярных областей и горячих источниках, на теле животных и растений и внутри организма. Особенно много их в почве — от 200-500 млн. до 2 млрд. и более особей в 1 г в зависимости от типа почвы.

Рис. 6. Бактерии: 1, 2 — бациллы; 3 — спирохеты; 4 — кокки; 5 — спириллы; 6 — вибрионы; 7 — стрептококки и диплококки; 8 — сарцины; 9 — нитевидные и 10 — жгутиковые формы
По форме и особенностям объединения клеток различают несколько морфологических групп бактерий: шаровидные (кокки), прямые палочковидные (бациллы), изогнутые (вибрионы) спирально изогнутые (спириллы) и др. Кокки, сцепленные попарно, получили название диплококки, соединенные в виде цепочки — стрептококки, в виде гроздей — стафилококки и др. Реже встречаются нитчатые формы (рис.6).
Прокариоты широко различаются по своим физиологическим свойствам и очень быстро делятся. Так, в течение 10-11 часов потомство одной единственной клетки в благоприятных условиях может достичь 4 млрд. особей. Прокариоты легко адаптируются к условиям окружающей среды, у них очень часты спонтанные мутации, а необычайное биохимическое многообразие способствует их повсеместному распространению на Земле.
Плазматическая мембрана (цитолемма) прокариот выполняет все свойственные функции: транспортную, защитную, разграничительную, рецепции, восприятия сигналов внешней (для клетки) среды, участия в иммунных процессах, обеспечения поверхностных свойств клетки. Кроме того, плазматические мембраны у них выполняют еще ряд важнейших функций: в них локализуются ферменты цепи переноса электронов и окислительного фосфорилирования, осуществляется синтез компонентов клеточной стенки и капсулы, выведение внеклеточных ферментов. У фотосинтезирующих организмов фотосинтез также осуществляется на мембране; на внутренней стороне мембраны расположены сайты связывания ДНК, каждая из дочерних молекул ДНК после репликации прикрепляется к одному из сайтов, в результате роста мембраны молекулы ДНК расходятся, после чего формируется перемычка, разделяющая клетку на две (рис. 7).
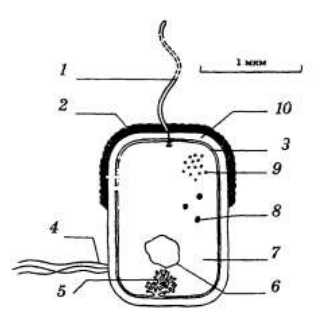
Рис.7. Строение бактериальной клетки: 1 — жгутик; 2 — капсула; 3 — мембрана; 4 — пили; 5 — мезосома; 6 — кольцевая ДНК; 7 — цитоплазма; 8 — запасные питательные вещества; 9 — рибосома; 10 — клеточная стенка
Общий принцип устройства мембран прокариот не отличается от эукариот, но в химическом составе имеются различия (отсутствует молекулы холестерина и некоторых других липидов, присущих мембранам эукариот). У некоторых микроорганизмов плазматическая мембрана впячивается внутрь клетки, образуя стопки плоских мешочков, связанных с цитолеммой. У цианобактерий и некоторых пурпурных бактерий имеется множество мембранных тилакоидов, связанных с мембраной и осуществляющих фотосинтез. Тилакоиды также являются производными цитолеммы, которые образуются в результате ее впячиваний. Некоторые прокариоты содержат газовые вакуоли (аэросомы). Все мембранные структуры прокариотической клетки непрерывны.
Клеточная стенка прокариот выполняет многочисленные функции, в первую очередь, формообразование и защита от осмотического шока. В различных клетках клеточная стенка составляет от 5 до 50% сухой массы клетки.
В соответствии с окраской, бактерии делят на две группы -грамположительные и грамотрицательные, что связано с особенностями строения их клеточных стенок. У грамположительных клеток цитолемма окружена толстой (20-80 нм) клеточной стенкой, состоящей из пептидогликана муреина, тейхоевых кислот и полисахаридов; у грамотрицательных — тонким (2-3 нм) слоем пептидогликана, покрытым наружной плазматической мембраной. Если обработать клетку лизоцимом или пенициллином, клеточная стенка разрушается. Грамположительная клетка превращается в протопласт, который весьма подвержен осмотическому шоку; грамотрицательная — в сферопласт, покрытый двумя мембранами и вследствие этого менее чувствительный к осмотическому шоку.
Капсулы, слизь, влагалища. Многие бактерии (например, пневмококки, клебсиеллы, некоторые клостридии и др.) и некоторые сине-зеленые водоросли кнаружи от клеточной стенки имеют слой сильно оводненного материала, образующего капсулу.
Капсулы связаны с клеточной стенкой, имеют толщину до 10 мкм. Капсулы обеспечивают устойчивость бактерий к ряду воздействий, например к фагоцитозу (повышая вирулентность бактерий), придают бактериям специфичность (в ней могут находиться антигены), предохраняют клетку от высыхания, механических повреждений, от действия вирусов, служат источником запасных питательных веществ, а также осуществляют связь между клетками и способствуют прикреплению к каким-либо поверхностям.
Подвижность прокариот. Прокариоты обладают жгутиками, расположение которых может быть монополярным, биполярным и перитрихиальным. По количеству жгутиков различают монотрихи (одна нить) и политрихи (пучок нитей). Скорость движения прокариот очень велика — от 1,6 до 12 мм/мин. Структура жгутика сложна и совершенно отлична от строения ресничек и жгутиков эукариот. Подвижные прокариоты совершают направленные движения по градиенту концентрации некоторых веществ — аттрактантов или против градиента концентрации других веществ — репеллентов. Такое поведение называется хемотаксисом. При движении жгутиков против часовой стрелки клетка движется в одном направлении, при движении жгутиков по часовой стрелке клетка кувыркается. Под влиянием аттрактантов клетка большую часть пути проделывает прямолинейно, под влиянием репеллентов клетка чаще кувыркается и удаляется. Помимо хемотаксиса, существуют и другие виды таксиса. Аэротаксис -аэробные бактерии устремляются к воздушной среде, а анаэробные, наоборот, скапливаются в центре культуры. Фототаксис — пурпурные бактерии перемещаются в зоны с наибольшей освещенностью» что связано с потребностью фототрофных бактерий в квантах света для получения энергии. Магнитотаксис — железосодержащие бактерии движутся в магнитном поле в направлении линий.
Фимбрии. Фимбрии, или пили, — это полые нити диаметром до 0,0015 мкм и длиной от 0,3-5 мкм, образованные белком полином. Они располагаются по периферии клетки в количестве 100-250. Фимбрии участвуют в процессах прикрепления бактерий к клеткам млекопитающих (например, к слизистой оболочке кишечника). У некоторых бактерий есть специализированные фимбрии, в частности F-фимбрии у кишечной палочки, участвующие в процессе конъюгации и могущие служить органом прикрепления бактериофагов.
Другие органеллы прокариот. Важными отличительными особенностями прокариот являются малое количество мембранных органелл и наличие множества рибосом (от 5 000 до 50 000 в клетке).
Внутриклеточные запасные вещества. В прокариотических клетках могут откладываться полисахариды, липиды, полифосфаты. Каждый вид прокариот накапливает один тип запасных веществ.
Покоящиеся формы. При неблагоприятных условиях внешней среды многие прокариоты образуют покоящиеся формы, способные сохранять жизнеспособность в течение длительного времени. К таким формам относят эндо- и экзоспоры, цисты, бактероиды, гетероцисты и др. Зрелая спора устойчива к различным неблагоприятным условиям среды, включая высокую температуру, радиацию, ультрафиолетовые лучи, химические агенты. Споры могут сохраняться в течение очень длительного времени (до 1000 лет) при нормальных условиях. Однако при температуре 100°С 90% спор бактерий гибнет через 11 минут; 99% спор актиномицетов погибает при температуре 75°С через 70 минут, а при высушивании они сохраняются до 15 лет. При улучшении условий внешней среды спора прорастает. Прорастание протекает в три этапа: активация, инициация и собственно прорастание. При прорастании спора теряет устойчивость к внешним факторам.
Питание бактерий. По типу питания бактерии делят на две группы: автотрофные и гетеротрофные. Автотрофные бактерии синтезируют органические вещества из неорганических. В зависимости оттого, какую энергию используют автотрофы для синтеза органических веществ, различают фото- (зеленые и пурпурные серобактерии) и хемосинтезирующие бактерии (нитрифицирующие, железобактерии, бесцветные серобактерии и др.). Гетеротрофные бактерии питаются готовыми органическими веществами отмерших остатков (сапротрофы) или живых растений, животных и человека (симбионты).
К сапротрофам относятся бактерии гниения и брожения. Первые расщепляют азотсодержащие соединения, вторые — углерод-содержащие. В обоих случаях выделяется энергия, необходимая для их жизнедеятельности.
Размножение. Бактерии размножаются путем простого бинарного деления клетки. Этому предшествует самоудвоение (репликация) молекулы ДНК. Почкование встречается как исключение.
У некоторых бактерий обнаружены упрощенные формы полового процесса. Например, у кишечной палочки половой процесс напоминает конъюгацию, при которой происходит передача части генетического материала из одной клетки в другую при их непосредственном контакте. После этого клетки разъединяются. Количество особей в результате полового процесса остается прежним, но происходит обмен наследственным материалом, т. е. осуществляется генетическая рекомбинация.
Спорообразование свойственно только небольшой группе бактерий, у которых известны два типа спор: эндогенные, образующиеся внутри клетки, и микроцисты, образующиеся из целой клетки. При образовании спор (микроцист) в бактериальной клетке уменьшается количество свободной воды, снижается ферментативная активность, протопласт сжимается и покрывается очень плотной оболочкой. Несмотря на постоянную гибель бактерий (поедание их простейшими, действие высоких и низких температур и других неблагоприятных факторов), эти примитивные организмы сохранились с древнейших времен благодаря способности к быстрому размножению (клетка может делиться через каждые 20-30 мин), образованию спор, чрезвычайно устойчивых к факторам внешней среды, и их повсеместному распространению.
Энергетический обмен. У микроорганизмов, как и у эукари-от, различают два типа дыхания: аэробное и анаэробное. Различают несколько групп микроорганизмов.
Строгие (облигатные) аэробы растут только при наличии воздуха, обладают набором ферментов для аэробного дыхания, производят полное окисление углеводов до С02 и Н20.
Строгие анаэробы могут развиваться лишь в бескислородной среде, обладают анаэробным дыханием.
Факультативные анаэробы способны развиваться как в бескислородных, так и в кислородных условиях. Процесс дыхания у них протекает в две фазы — сначала фаза анаэробного роста, а затем потребление кислорода и более глубокое расщепление углеводов. Они обладают обоими наборами ферментов.
Микроаэрофилы развиваются при низких концентрациях кислорода — до 1%. К ним относят спирохеты, актиномицеты.
Аэротолерантные микробы могут развиваться при пониженных концентрациях кислорода — до 10%. Примером могут служить клостридии.
Капнические микробы требуют повышенного содержания углекислоты (например, Brucella abortus).
Энергетический обмен у прокариот в основном включает в себя те же этапы, что и у эукариот.
Генетический аппарат прокариот. Одним из отличий прокариот является отсутствие у них оформленного ядра. Вместо него имеется нуклеоид. Величина генома у бактерий составляет от 0,8х106 до 8х106 пар нуклеотидов. Помимо хромосомы, у бактерий имеются плазмиды -суперспирализованные молекулы двухнитчатой ДНК, ковалентно замкнутые в кольцевую структуру. Бактериальные клетки могут содержать несколько плазмид. Плазмиды способны реплицироваться независимо от хромосомы. Плазмиды могут находиться в бактериальной клетке в двух состояниях — автономном (в цитоплазме) и интегрированном (встраиваются структуру бактериальной хромосомы и реплицируются вместе с ней). Плазмиды способны переноситься из клетки в клетку. Часто они контролируют у бактерий определенные свойства. К плазмидам относят также профагов — стадия существования умеренных бактериофагов. С плазмидами связывают патогенность ряда бактерий и их отдельных штаммов.
Боррелии. Бактерии относятся к порядку Spirochaetales, семейству Spirochaetaceae, роду Borrelia. Свое родовое название они получили в честь французского микробиолога A.Borrel, изучавшего в начале ХХ века эту разновидность спирохет. Все известные виды рода Borrelia морфологически весьма сходны. Это грамотрицательные спирохеты, относительно легко окрашиваются анилиновыми красителями, что отличает их от других родов спирохет. Другой отличительной особенностью боррелий является отсутствие у них митохондрий и ундулирующей мембраны. Длина микробной клетки — от 10 до 30 мкм, поперечный размер составляет 0,20-0,25 мкм. По форме представляют собой извитую, лево- или правовращающуюся спираль.
В настоящее время по отличиям в нуклеотидных последовательностях ДНК различают более 10 (13) геновидов боррелий, относящихся к комплексу B.burgdorferi sensu lato, представители которого неравномерно распределены в пределах мирового нозоареала этой инфекции. Не все боррелии данного комплекса являются патогенными для человека. Доказана патогенность 3 геновидов: B.burgdorferi sensu stricto, B.garinii и B.afzelii. Все эти геновиды имеют распространение на территории России (преимущественно B.garinii и B.afzelii).
Боррелии in vitro культивируются только в среде обогащенной аминокислотами, витаминами, альбумином бычьей и кроличьей плазмы и другими веществами (модифицированная среда BSK-II). Температурный оптимум культивирования составляет 30-34оС. Боррелии отличаются относительно длительным и непостоянным периодом роста на питательных средах в анаэробных условиях, который обычно составляет 7-20 часов. При достижении числа микробных клеток 106 — 108 в мл возможно осуществление дальнейших лабораторных исследований. Боррелии выращенные в среде культивирования хорошо сохраняются при низких температурах (-70°-90°С) до нескольких лет, не теряя своих биологических свойств. Формалин, фенол, этиловый спирт и другие дезинфицирующие вещества, а также ультрафиолетовое излучение обладают инактивирующим действием на боррелии.
Отсутствие у боррелий генов кодирования синтеза амино- и жирных кислот, ко-факторов и нуклеотидов объясняет столь высокие требования к условиям их культивирования с использованием сывороточных добавок. Кроме того, поскольку боррелии хемоорганотрофные анаэробы, то они должны получать энергию для синтеза АТФ от субстрата фосфорилирования, который имеется только в цитоплазме других клеток, т.к. у боррелий нет генов, кодирующих дыхательную цепь фосфорилирования. Боррелии также испытывают недостаток других ферментов, необходимых для цикла трикарбоновых кислот и фосфорилирования. Таким образом, способность их собственных компонентов метаболизма настолько ограничена, что для своего существования они должны иметь организм хозяина (причем с преимущественным внутриклеточным персистированием), который гарантирует доступность питательных готовых веществ.
Боррелии имеют одну линейную хромосому и, кроме того, еще до 20 кольцевых и линейных плазмид, известных еще под названием -микрохромосомы.
Экстрахромосомный (плазмидный) феномен генного кодирования не является исключительным свойством боррелий, подобное наблюдается и у некоторых других представителей прокариот. Однако тот факт, что боррелии могут иметь довольно большое количество линейных и кольцевых плазмид выделяет их среди других микроорганизмов. Следующей особенностью боррелий является то, что плазмиды обнаруживаются преимущественно при первичной изоляции штаммов в культуре, последующие пересевы приводят к уменьшению их числа и даже полной элиминации, в связи с чем, изменяется и антигенный профиль возбудителя, приводящий к полной потере инвазивности.
Белковый (антигенный) спектр боррелий весьма вариабелен и различен не только у боррелий разных геновидов, но и среди изолятов одного геновида.
Боррелии имеют группы антигенов: поверхностные (OspA, OspB, OspD, OspE и OspF), жгутиковый и цитоплазматический. Поверхностные антигены характеризуются вариабельностью. Наибольшая их гетерогенность обнаруживается у B.garinii (по OspC — 13 вариантов, по OspA — 7) и B.afzelii (по OspC — 8, по OspA — 2). Штаммы B.burgdorferi s.s. отличаются относительным постоянством спектра поверхностных белков. Белки, находящиеся на внешней оболочке, определяют видовую принадлежность возбудителя и являются основными иммуногенами. Многие антигенные детерминанты внешней оболочки сходны с таковыми у боррелий других видов и даже некоторых бактерий. Этим объясняется возможность перекреста в иммунологических реакциях.
В настоящее время установлены и достаточно хорошо изучены и некоторые другие мембранные антигены боррелий: p83/100, Oms66 (p66), BmpA (p39), HSP60 (p60), p18, OspD, OspE, OspF.
Генетическая информация большинства из них находится в плазмидах, поэтому, потеря отдельных плазмид, например, при длительной персистенции боррелий в организме человека, обязательно влечет за собой и изменение структуры поверхностных антигенов, а значит и иммунного ответа.
- Подробности
- Раздел: Паразитология и природная очаговость болезней
zoovet.info
Бактерии, эукариоты, простейшие – почему они такие разные?
Бактерии относятся к доядерным одноклеточным микроорганизмам прокариотам, то есть у них отсутствует ядерная белковая оболочка – упаковка для ДНК. Также их структура более упрощена по сравнению с клетками животных и растений. Основной тип питания – это фотосинтез (при помощи энергии света) или хемосинтез (окисление веществ). К прокариотам относятся также археи, сине-зеленые водоросли.
Эукариоты – это надцарство живых организмов, клетки которых имеют ядро, и его оболочка четко оформлена. С греческого языка термин переводится как «хорошее ядро», вот почему выбрано именно это название.
К этому надцарству принадлежат растения, животные, грибы, простейшие, грибоподобные организмы слизевики, водоросли.
Существует теория, что древняя цианобактерия около 2,5 млрд. лет назад была захвачена клеткой – предшественником эукариота, что обусловило появление абсолютно новых микроорганизмов. Некоторые отдельные органеллы эукариотов (к примеру, митохондрии и пластиды) очень похожи на бактерий по строению и особенностям жизнедеятельности. Они также размножаются делением, имеют собственный генетический аппарат.
Почему эукариоты не являются бактериями или археями
От бактерий (прокариотов) и архей основным отличием эукариотов является расположение генетического аппарата в окружении двойной мембраны, защищенного крепкой оболочкой ядра. Встречаются многоядерные организмы. У них линейная ДНК, связанная с гистонами – белками, в которые упакованы нити. У бактерий же ДНК кольцевидная, не связанная гистонами.
Клетка насчитывает десятки постоянных структур – ее органоидов, обеспечивающих жизнедеятельность, каждый из которых отделен мембраной одной или несколькими. Это достаточно редко встречается у прокариотов.
Наличие митохондрий (органеллы в форме сферы или эллипса, отделенные двумя мембранами) обеспечивает выработку энергии за счет окисления органических соединений, которая является источником функционирования организма. Простейшее-паразит трипаносома имеет одну большую митохондрию, амеба – около 0,5 млн., у прокариотов их нет вообще.
Наличие пластид, которые могут состоять из 4 мембран, также существенно отличает прокариотов от эукариотов. Пластиды окружены внешней и внутренней мембраной и выполняют:
- функции фотосинтеза,
- синтез аминокислот, пуринов, абсцизовой кислоты и других важных соединений.
Пластиды обеспечивают запасы липидов, крахмала, железа.
Размеры эукариотов в тысячи раз больше прокариотов. Вот почему для сохранения жизни им необходимо поглощать большое количество белка в качестве питания. Это обусловило появление хищных организмов.
Особенности строения
Стандартная клетка состоит из следующих структур:
- ядро,
- рибосома,
- везикула,
- шероховатый эндоплазматический ретикулум,
- аппарат Гольджи,
- гладкий эндоплазматический ретикулум,
- митохондрия,
- вакуоль,
- гиалоплазма,
- лизосома,
- центросома,
- меланосома,
- реснички, жгутики,
- клеточная стенка.
Ядро содержит ядрышко, которое не имеет мембранной оболочки. Оно хорошо различимо под электронным микроскопом. В ядрышке происходит синтез РНК. Ядро обеспечивает хранения ДНК – наследственной информации, ее передачу, реализацию, воспроизводство.
Рибосома, являясь органоидом, имеет форму сферы, осуществляет трансляцию (синтез белка из аминокислот). Рибосомы бывают большие и маленькие.
Строение клетки эукариот
Везикула – небольшой органоид, отделенный мембраной, образующий внутриклеточную сумку для транспортировки или преобразования питательных веществ, хранения ферментов.
Шероховатый (гранулярный) эндоплазматический ретикулум состоит из разветвлений, характеризуется наличием пузырьков, трубочек и полостей. Он окружен мембранной оболочкой. На его поверхности содержатся рибосомы, осуществляющие синтез белков.
Аппарат Гольджи — структура, состоящая из мембран и «цистерн», помогающая выведению из гранулярного эндоплазматического ретикулума веществ. По внешнему виду напоминает трубки, собранные в стопки. В цистернах происходит созревание белков, в каждом отделе содержится свой набор ферментов. Везикулы, отделяясь от ретикулума, непрерывно присоединяются к аппарату Гольджи. Когда белок готов переместиться, пузырьки отсоединяются и доставляются к необходимой органелле. Аппарат Гольджи сортирует вещества, оправляя некоторые из них к плазматической мембране, другие к лизосомам.
Гладкий (агранулярный) эндоплазматический ретикулум не имеет рибосом. Отвечает за процессы метаболизма. Осуществляет синтез липидов, жирных кислот, стероидов. Ткани печени и надпочечников состоят из гладкого эндоплазматического ретикулума.
Митохондрии – органоиды, окисляющие органические соединения, используя энергию для обеспечения жизни всего организма. Могут различаться по формам, количество, содержащееся в одной клетке, может варьировать от одной митохондрии до сотен тысяч. В ней содержится кольцевая спиральная молекула ДНК.
Вакуоли развиваются из пузырьков мембран. Их имеют не все эукариоты. Выполняют функцию накопления воды, выводят продукты распада. Бывают пищеварительными, пульсирующими.
Гиалоплазма – это внутриклеточная жидкость.
Лизосома – органоид, вид везикул, окруженный мембраной, содержащий в себе ферменты. Выполняет функцию переваривания молекул, посредством секреции. Прокариоты не имеют лизосом.
Центросома регулирует процессы клеточного деления, образования трубочек, являясь немембранным органоидом. Участвует в формировании жгутиков, ресничек.
Меланосома присутствует у животных, содержит светопоглощающие пигменты, в частности, меланин.
Реснички – это тоненькие волоски на поверхности клеточной стенки, покрытые мембраной, являющиеся рецепторами. Они есть у инфузорий, губок, реснитчатых червей. Их имеют клетки эпителия кишечника, дыхательные пути – бронхи, мозговые желудочки, евстахиева труба.
Жгутики могут быть также у прокариотов. У бактерий они намного тоньше, короче, не могут изгибаться. Жгутики эукариотов длиннее ресничек, хотя схожи с ними по структуре. У архебактерий жгутики несколько тоньше, отличаются строением.
Клеточная стенка, в первую очередь, обеспечивает защиту всех внутренних структур от внешних факторов, а также выполняет транспортировку веществ. Состоит из муреина, структура которого влияет на степень окрашивания ее по методу Грама. Некоторые бактерии, водоросли, грибы, археи тоже имеют клеточную стенку. Также бактерии могут образовывать капсулу – слизистую структуру из полисахаридов, большого количества воды вокруг стенки.
Жизнь и питание эукариотов
Цикл жизни эукариотов разделен на две последующие фазы:
- гаплофаза,
- диплофаза.
Происходит слияние двух галоплоидных (с одним набором хромосом) клеток и их ядер в одну общую, имеющую два (диплоидных) набора хромосом. Через некоторое время клетки вновь становятся галоплоидными, разделяясь. Такой способ совершенно нехарактерен для прокариотов.
Отличием бактерий, архей и эукариотов является способность последних к эндоцитозу – захвату других клеток и помещению их в специальные сумки (везикулы), в которых путем ферментации происходит «переваривание» пищи до консистенции, способной проникнуть сквозь клеточную мембрану.
Некоторые способны к фагоцитозу (с греческого «пожирание»). Они могут захватывать твердые частицы (вирусы, бактерии), переваривать их, осуществляя таким образом питание.
Также эукариоты умеют впитывать жидкость. Пиноцитоз – способность всех эукариотических клеток поглощать молекулы воды, других жидких веществ, утоляя свою потребность в питье.
Особенности строения, различие в течение процессов, отвечающих за жизнедеятельность клеток, а также размеры, наличие органов, выполняющих определенные функции – все это существенно отличает эукариотов от бактерий. Вот почему они не являются бактериями, а представляют собой отдельный вид микроорганизмов.
probakterii.ru
Leave A Comment