
Естественный отбор. Большая российская энциклопедия
Природные процессы, явления в живых организмах
Есте́ственный отбо́р, процесс избирательного выживания и дифференциального размножения организмов, основной движущий фактор их эволюции. Естественный отбор носит вероятностный характер, действует на основе мутационного процесса и существующего генофонда, влияет на частоту распространения генов и их комбинаций, способствует уменьшению негативного действия мутаций и образованию механизмов защиты от их вредного действия, определяя тем самым темпы и направления эволюции.
Историческая справка
Идеи о существовании естественного отбора высказывались с начала 19 в. разными английскими натуралистами (в том числе А. Уоллесом). Но только Ч. Дарвин (1842, 1859) оценил его как главный фактор эволюции. По Дарвину, естественный отбор является результатом борьбы за существование; даже незначительные наследуемые различия между особями одного вида могут дать преимущества в этой борьбе, которая обусловлена тенденцией организмов к высокой интенсивности размножения (в геометрической прогрессии) и невозможности сохранения всего потомства вследствие ограниченности природных ресурсов. Гибель подавляющего числа особей в каждом поколении неизбежно ведёт к естественному отбору – «выживанию наиболее приспособленного» к данным условиям. В результате суммирования полезных изменений в течение многих поколений формируются новые адаптации и в конечном счёте возникают новые виды. Рассуждения о действии естественного отбора Дарвин строил преимущественно на обобщении опыта доместикации животных и растений по аналогии с искусственным отбором, подчёркивая, однако, что в отличие от проводимой человеком селекции естественный отбор определяется взаимодействием организмов с условиями окружающей среды и не имеет определённой цели.
Гибель подавляющего числа особей в каждом поколении неизбежно ведёт к естественному отбору – «выживанию наиболее приспособленного» к данным условиям. В результате суммирования полезных изменений в течение многих поколений формируются новые адаптации и в конечном счёте возникают новые виды. Рассуждения о действии естественного отбора Дарвин строил преимущественно на обобщении опыта доместикации животных и растений по аналогии с искусственным отбором, подчёркивая, однако, что в отличие от проводимой человеком селекции естественный отбор определяется взаимодействием организмов с условиями окружающей среды и не имеет определённой цели.
Систематическое исследование естественного отбора, расширение и совершенствование методик его изучения началось с конца 19 в. Использование методов биометрии позволило установить статистически значимые различия между выжившими и погибшими организмами при изменении условий среды. Благодаря разработкам Р. Фишера, Дж. Холдейна, С. Райта и С. С. Четверикова, осуществивших синтез классического дарвинизма и генетики, появилась возможность приступить к экспериментальному изучению генетических основ естественного отбора. Обследованные природные популяции оказались буквально насыщены мутациями, многие из которых становились полезными при изменениях условий существования или при комбинации с другими мутациями. Было установлено, что мутационный процесс и свободное скрещивание (панмиксия) обеспечивают генетическую разнородность популяций и уникальность особей, имеющих разные шансы на выживание; это обусловливает высокую интенсивность и эффективность естественного отбора. Кроме того, стало очевидным, что естественный отбор имеет дело не с единичными признаками, а с целыми организмами и что генетическая сущность естественного отбора заключается в неслучайном (дифференцированном) сохранении в популяции определённых генотипов, избирательно передающихся следующим поколениям.
Обследованные природные популяции оказались буквально насыщены мутациями, многие из которых становились полезными при изменениях условий существования или при комбинации с другими мутациями. Было установлено, что мутационный процесс и свободное скрещивание (панмиксия) обеспечивают генетическую разнородность популяций и уникальность особей, имеющих разные шансы на выживание; это обусловливает высокую интенсивность и эффективность естественного отбора. Кроме того, стало очевидным, что естественный отбор имеет дело не с единичными признаками, а с целыми организмами и что генетическая сущность естественного отбора заключается в неслучайном (дифференцированном) сохранении в популяции определённых генотипов, избирательно передающихся следующим поколениям.
Роль естественного отбора в эволюционном процессе
Естественный отбор носит вероятностный характер, действует на основе мутационного процесса и существующего генофонда, влияет на частоту распространения генов и их комбинаций, способствует уменьшению негативного действия мутаций и образованию механизмов защиты от их вредного действия, определяя тем самым темпы и направления эволюции. Под контролем естественного отбора находятся не только разнообразные признаки, но и сами факторы эволюции, например интенсивность и характер мутабельности, аппарат наследственности (отсюда понятие «эволюция эволюции»). При отсутствии же естественного отбора происходит снижение или утрата приспособленности организмов из-за накопления нежелательных мутаций, что проявляется в возрастании генетического груза, в том числе в популяциях современного человека.
Под контролем естественного отбора находятся не только разнообразные признаки, но и сами факторы эволюции, например интенсивность и характер мутабельности, аппарат наследственности (отсюда понятие «эволюция эволюции»). При отсутствии же естественного отбора происходит снижение или утрата приспособленности организмов из-за накопления нежелательных мутаций, что проявляется в возрастании генетического груза, в том числе в популяциях современного человека.
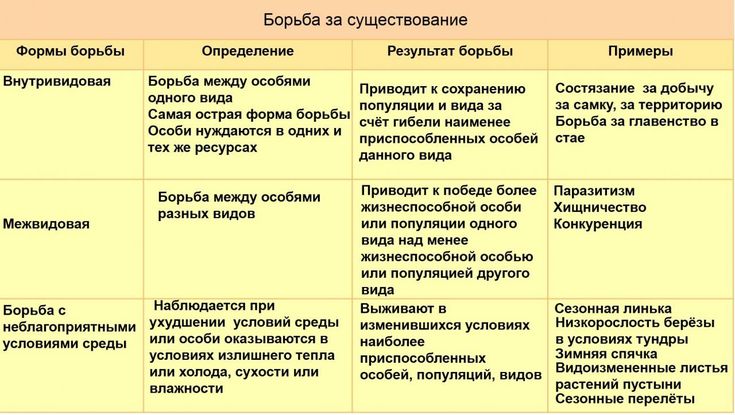
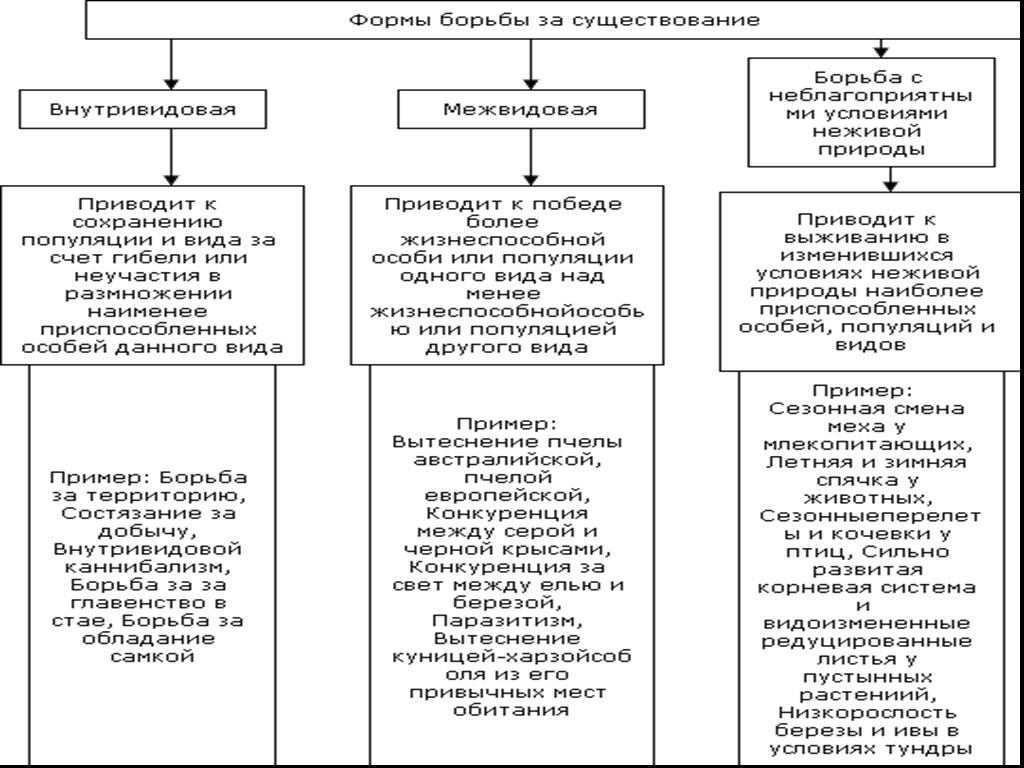
Формы естественного отбора
Выделяют более 30 форм естественного отбора; ни одна из них не существует в чистом виде, а скорее характеризует тенденцию действия отбора в конкретной экологической ситуации. Так, движущий отбор способствует сохранению определённого отклонения от прежней нормы и приводит к выработке новых приспособлений через направленную перестройку всего генофонда популяций, а также генотипов и фенотипов особей. Он может вести к доминированию одной (или нескольких) ранее существовавшей формы над другими. Классическим примером его действия стало преобладание в промышленных районах темноокрашенных форм бабочки берёзовая пяденица, незаметных для птиц на загрязнённых копотью стволах деревьев (до середины 19 в. встречалась только светлая форма, имитировавшая пятна лишайников на светлых стволах берёзы). Быстрое привыкание различных видов насекомых и грызунов к ядам, возникновение резистентности микроорганизмов к антибиотикам свидетельствуют о том, что давления движущего отбора в природных популяциях достаточно для того, чтобы обеспечить быстрый адаптивный ответ на резкие изменения среды. Как правило, селекция по одному признаку влечёт за собой целый ряд преобразований. Например, длительный отбор на содержание белка или масла в зёрнах кукурузы сопровождается изменениями форм зёрен, размеров початков, их расположения над уровнем почвы и др.
встречалась только светлая форма, имитировавшая пятна лишайников на светлых стволах берёзы). Быстрое привыкание различных видов насекомых и грызунов к ядам, возникновение резистентности микроорганизмов к антибиотикам свидетельствуют о том, что давления движущего отбора в природных популяциях достаточно для того, чтобы обеспечить быстрый адаптивный ответ на резкие изменения среды. Как правило, селекция по одному признаку влечёт за собой целый ряд преобразований. Например, длительный отбор на содержание белка или масла в зёрнах кукурузы сопровождается изменениями форм зёрен, размеров початков, их расположения над уровнем почвы и др.
Результатом действия движущего отбора в филогенезе крупных таксонов является ортоселекция, примером которой служит установленная В. О. Ковалевским направленная эволюция конечности предков лошади (от пятипалости к однопалости), протекавшая миллионы лет и обеспечившая увеличение скорости и экономичности бега.
Дизруптивный, или разрывающий, отбор благоприятствует сохранению крайних отклонений и ведёт к увеличению полиморфизма. Он проявляется в тех случаях, когда ни одна из внутривидовых форм с разными генотипами не получает абсолютного преимущества в борьбе за существование из-за разнообразия условий, одновременно встречающихся на одной территории; при этом прежде всего элиминируются особи со средним или промежуточным характером признаков. Ещё в начале 20 в. российский ботаник Н. В. Цингер показал, что большой погремок (Alectoroleophus major), цветущий и плодоносящий на некошеных лугах в течение всего лета, на скашиваемых лугах образует две расы: ранневесеннюю, успевающую принести семена до начала покоса, и позднеосеннюю – низкие растения, не повреждаемые при покосе, а затем быстро зацветающие и успевающие дать семена до начала морозов. Другим примером полиморфизма служит различие в окраске раковин у земляной улитки (Capaсea nemoralis), являющейся кормом для птиц: в густых буковых лесах, где в течение всего года сохраняется подстилка из красно-бурого опада, обычны особи с коричневой и розовой окраской; на лугах с жёлтой подстилкой преобладают улитки с жёлтой окраской.
Он проявляется в тех случаях, когда ни одна из внутривидовых форм с разными генотипами не получает абсолютного преимущества в борьбе за существование из-за разнообразия условий, одновременно встречающихся на одной территории; при этом прежде всего элиминируются особи со средним или промежуточным характером признаков. Ещё в начале 20 в. российский ботаник Н. В. Цингер показал, что большой погремок (Alectoroleophus major), цветущий и плодоносящий на некошеных лугах в течение всего лета, на скашиваемых лугах образует две расы: ранневесеннюю, успевающую принести семена до начала покоса, и позднеосеннюю – низкие растения, не повреждаемые при покосе, а затем быстро зацветающие и успевающие дать семена до начала морозов. Другим примером полиморфизма служит различие в окраске раковин у земляной улитки (Capaсea nemoralis), являющейся кормом для птиц: в густых буковых лесах, где в течение всего года сохраняется подстилка из красно-бурого опада, обычны особи с коричневой и розовой окраской; на лугах с жёлтой подстилкой преобладают улитки с жёлтой окраской. В смешанных же лиственных лесах, где характер фона меняется с наступлением нового сезона, ранней весной доминируют улитки с бурой и розовой окраской, а летом – с жёлтой. Дарвиновы вьюрки (Geospizinae) на островах Галапагос (классический пример адаптивной радиации) – конечный результат длительного дизруптивного отбора, приведшего к образованию десятков близкородственных видов.
В смешанных же лиственных лесах, где характер фона меняется с наступлением нового сезона, ранней весной доминируют улитки с бурой и розовой окраской, а летом – с жёлтой. Дарвиновы вьюрки (Geospizinae) на островах Галапагос (классический пример адаптивной радиации) – конечный результат длительного дизруптивного отбора, приведшего к образованию десятков близкородственных видов.
Если указанные формы естественного отбора приводят к изменению и фенотипической, и генетической структуры популяций, то впервые описанный И. И. Шмальгаузеном (1938) стабилизирующий отбор сохраняет в популяции среднее значение признаков (норму) и не пропускает в следующее поколение геномы особей, наиболее отклоняющихся от этой нормы. Он направлен на поддержание и повышение устойчивости в популяции среднего, ранее сложившегося фенотипа. Известно, например, что во время снежных бурь выживают птицы, которые по многим признакам (длина крыла, клюва, масса тела и т. д.) приближаются к средней норме, а отклонившиеся от этой нормы особи погибают. Размеры и форма цветков у растений, опыляемых насекомыми, более устойчивы, чем у растений, опыляемых ветром, что обусловлено сопряжённой эволюцией растений и их опылителей, «выбраковкой» уклонившихся от нормы форм (например, шмель не может проникнуть в слишком узкий венчик цветка, а хоботок бабочки не касается слишком коротких тычинок у растений с длинным венчиком). Благодаря стабилизирующему отбору при внешнем неизменном фенотипе могут идти существенные генетические изменения, обеспечивающие независимость развития адаптаций от колеблющихся условий среды. Одним из результатов действия стабилизирующего отбора можно считать «биохимическую универсальность» жизни на Земле.
Размеры и форма цветков у растений, опыляемых насекомыми, более устойчивы, чем у растений, опыляемых ветром, что обусловлено сопряжённой эволюцией растений и их опылителей, «выбраковкой» уклонившихся от нормы форм (например, шмель не может проникнуть в слишком узкий венчик цветка, а хоботок бабочки не касается слишком коротких тычинок у растений с длинным венчиком). Благодаря стабилизирующему отбору при внешнем неизменном фенотипе могут идти существенные генетические изменения, обеспечивающие независимость развития адаптаций от колеблющихся условий среды. Одним из результатов действия стабилизирующего отбора можно считать «биохимическую универсальность» жизни на Земле.
Дестабилизирующий отбор (название предложено Д. К. Беляевым, 1970) ведёт к резкому нарушению систем регуляции онтогенеза, вскрытию мобилизационного резерва и росту фенотипической изменчивости при интенсивном отборе в каком-либо определённом направлении. Например, отбор на снижение агрессивности хищных зверей в неволе через перестройку нейрогуморальной системы ведёт к дестабилизации цикла размножения, сдвигам в сроках линьки, изменениям в положении хвоста, ушей, в окраске и др.
Обнаружены гены, которые могут быть летальными или снижать жизнеспособность организмов в гомозиготном состоянии, а в гетерозиготном, напротив, повышать экологическую пластичность и другие показатели. В этом случае можно говорить о так называемом сбалансированном отборе, обеспечивающем поддержание генетического разнообразия с определённым соотношением частот аллелей. Примером его действия может служить повышение устойчивости у больных серповидно-клеточной анемией (гетерозиготных по гену гемоглобина S) к заражению различными штаммами малярийного плазмодия.
Важным шагом в преодолении стремления все признаки организмов объяснять действием естественного отбора стала концепция нейтральной эволюции, согласно которой часть изменений на уровне белков и нуклеиновых кислот происходит путём фиксации адаптивно нейтральных или почти нейтральных мутаций. Возможен отбор видов, возникающих в периферийных популяциях «внезапно» с геохронологической точки зрения.
Ещё раньше было доказано, что катастрофический отбор, при котором в период резких изменений среды выживает небольшое число особей и даже единственный организм, может стать основой формирования нового вида за счёт хромосомной перестройки и смены экологической ниши. Так, образование ксерофитного, эндемичного вида Clarkia lingulata в горах Сьерра-Невада в Калифорнии объясняют сильной засухой, вызвавшей массовую гибель растений, принявшую катастрофический характер в периферийных популяциях.
Так, образование ксерофитного, эндемичного вида Clarkia lingulata в горах Сьерра-Невада в Калифорнии объясняют сильной засухой, вызвавшей массовую гибель растений, принявшую катастрофический характер в периферийных популяциях.
Естественный отбор, затрагивающий вторичные половые признаки особей, называют половым (например, яркая брачная окраска самцов у многих видов рыб и птиц, зазывающие крики, специфические запахи, сильно развитые орудия для турнирного боя у млекопитающих). Эти признаки полезны, так как повышают возможность участия их носителей в воспроизведении потомства. В половом отборе наибольшую активность проявляют самцы, что выгодно для вида в целом, так как самки остаются в большей безопасности в период размножения.
Выделяют также групповой отбор, способствующий сохранению признаков, полезных семье, стае, колонии. Его частным случаем у колониальных насекомых является отбор сородичей, при котором стерильные касты (рабочие, солдаты и др.) обеспечивают (нередко ценой собственной жизни) выживание плодовитых особей (маток) и личинок и тем самым сохранение всей колонии.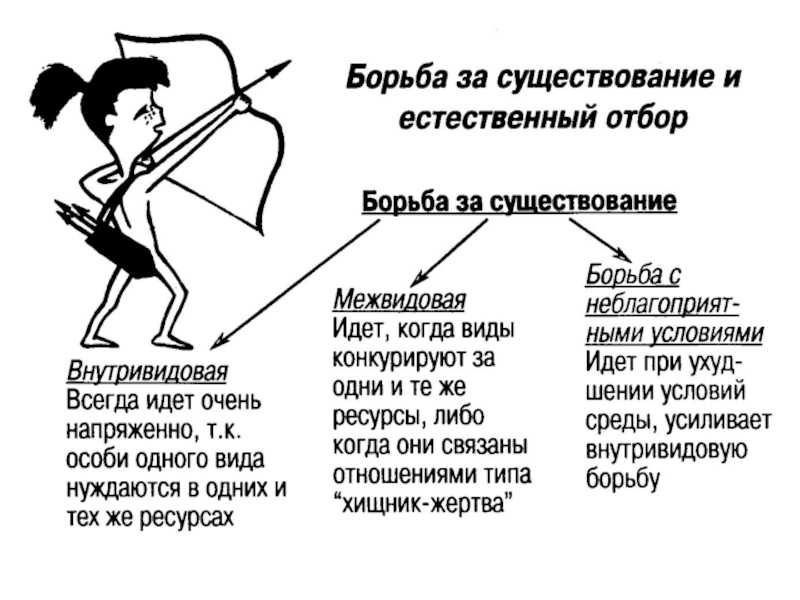 Альтруистическое поведение родителей, притворяющихся ранеными, чтобы увести хищника от своих детей, грозит гибелью имитатору, но в целом повышает шансы на выживание его потомства.
Альтруистическое поведение родителей, притворяющихся ранеными, чтобы увести хищника от своих детей, грозит гибелью имитатору, но в целом повышает шансы на выживание его потомства.
Критика существующих концепций
Хотя представления о ведущей роли естественного отбора в эволюции получили подтверждение во множестве экспериментов, до сих пор они подвергаются критике, исходящей из представления о невозможности образования организмов в результате случайной комбинации мутаций. При этом игнорируется тот факт, что каждый акт естественного отбора совершается на базе предыдущих результатов его же действия, которые, в свою очередь, предопределяют формы, интенсивность и направления естественного отбора, а значит, пути и закономерности эволюции.
Колчинский Эдуард Израилевич. Первая публикация: Большая российская энциклопедия, 2007.Дата публикации: 10 июня 2022 г. в 11:17 (GMT+3)
ЕСТЕСТВЕННЫЙ ОТБОР • Большая российская энциклопедия
ЕСТЕ́СТВЕННЫЙ ОТБО́Р, процесс избирательного выживания и дифференциального размножения организмов, основной движущий фактор их эволюции. Идеи о существовании Е. о. высказывались с нач. 19 в. разными англ. натуралистами (в т. ч. А. Уоллесом). Но только Ч. Дарвин (1842, 1859) оценил его как гл. фактор эволюции. По Дарвину, Е. о. является результатом борьбы за существование; даже незначит. наследуемые различия между особями одного вида могут дать преимущества в этой борьбе, которая обусловлена тенденцией организмов к высокой интенсивности размножения (в геометрич. прогрессии) и невозможности сохранения всего потомства вследствие ограниченности природных ресурсов. Гибель подавляющего числа особей в каждом поколении неизбежно ведёт к Е. о. – «выживанию наиболее приспособленного» к данным условиям. В результате суммирования полезных изменений в течение мн. поколений формируются новые адаптации и в конечном счёте возникают новые виды. Рассуждения о действии Е.
Идеи о существовании Е. о. высказывались с нач. 19 в. разными англ. натуралистами (в т. ч. А. Уоллесом). Но только Ч. Дарвин (1842, 1859) оценил его как гл. фактор эволюции. По Дарвину, Е. о. является результатом борьбы за существование; даже незначит. наследуемые различия между особями одного вида могут дать преимущества в этой борьбе, которая обусловлена тенденцией организмов к высокой интенсивности размножения (в геометрич. прогрессии) и невозможности сохранения всего потомства вследствие ограниченности природных ресурсов. Гибель подавляющего числа особей в каждом поколении неизбежно ведёт к Е. о. – «выживанию наиболее приспособленного» к данным условиям. В результате суммирования полезных изменений в течение мн. поколений формируются новые адаптации и в конечном счёте возникают новые виды. Рассуждения о действии Е. о. Дарвин строил преим. на обобщении опыта доместикации животных и растений по аналогии с искусственным отбором, подчёркивая, однако, что в отличие от проводимой человеком селекции Е. о. определяется взаимодействием организмов с условиями окружающей среды и не имеет определённой цели.
о. Дарвин строил преим. на обобщении опыта доместикации животных и растений по аналогии с искусственным отбором, подчёркивая, однако, что в отличие от проводимой человеком селекции Е. о. определяется взаимодействием организмов с условиями окружающей среды и не имеет определённой цели.
Систематич. исследование Е. о., расширение и совершенствование методик его изучения началось с кон. 19 в. Использование методов биометрии позволило установить статистически значимые различия между выжившими и погибшими организмами при изменении условий среды. Благодаря разработкам Р. Фишера, Дж. Холдейна, С. Райта и С. С. Четверикова, осуществивших синтез классич. дарвинизма и генетики, появилась возможность приступить к эксперим. изучению генетич. основ Е. о. Обследованные природные популяции оказались буквально насыщены мутациями, многие из которых становились полезными при изменениях условий существования или при комбинации с др. мутациями. Было установлено, что мутационный процесс и свободное скрещивание (панмиксия) обеспечивают генетич. разнородность популяций и уникальность особей, имеющих разные шансы на выживание; это обусловливает высокую интенсивность и эффективность Е. о. Кроме того, стало очевидным, что Е. о. имеет дело не с единичными признаками, а с целыми организмами и что генетич. сущность Е. о. заключается в неслучайном (дифференцированном) сохранении в популяции определённых генотипов, избирательно передающихся следующим поколениям. Е. о. носит вероятностный характер, действует на основе мутационного процесса и существующего генофонда, влияет на частоту распространения генов и их комбинаций, способствует уменьшению негативного действия мутаций и образованию механизмов защиты от их вредного действия, определяя тем самым темпы и направления эволюции.
мутациями. Было установлено, что мутационный процесс и свободное скрещивание (панмиксия) обеспечивают генетич. разнородность популяций и уникальность особей, имеющих разные шансы на выживание; это обусловливает высокую интенсивность и эффективность Е. о. Кроме того, стало очевидным, что Е. о. имеет дело не с единичными признаками, а с целыми организмами и что генетич. сущность Е. о. заключается в неслучайном (дифференцированном) сохранении в популяции определённых генотипов, избирательно передающихся следующим поколениям. Е. о. носит вероятностный характер, действует на основе мутационного процесса и существующего генофонда, влияет на частоту распространения генов и их комбинаций, способствует уменьшению негативного действия мутаций и образованию механизмов защиты от их вредного действия, определяя тем самым темпы и направления эволюции. Под контролем Е. о. находятся не только разнообразные признаки, но и сами факторы эволюции, напр. интенсивность и характер мутабельности, аппарат наследственности (отсюда понятие «эволюция эволюции»). При отсутствии же Е. о. происходит снижение или утрата приспособленности организмов из-за накопления нежелательных мутаций, что проявляется в возрастании генетич. груза, в т. ч. в популяциях совр. человека.
Под контролем Е. о. находятся не только разнообразные признаки, но и сами факторы эволюции, напр. интенсивность и характер мутабельности, аппарат наследственности (отсюда понятие «эволюция эволюции»). При отсутствии же Е. о. происходит снижение или утрата приспособленности организмов из-за накопления нежелательных мутаций, что проявляется в возрастании генетич. груза, в т. ч. в популяциях совр. человека.
Выделяют более 30 форм Е. о.; ни одна из них не существует в чистом виде, а скорее характеризует тенденцию действия отбора в конкретной экологич. ситуации. Так, движущий отбор способствует сохранению определённого отклонения от прежней нормы и приводит к выработке новых приспособлений через направленную перестройку всего генофонда популяций, а также генотипов и фенотипов особей. Он может вести к доминированию одной (или нескольких) ранее существовавшей формы над другими. Классич. примером его действия стало преобладание в пром. районах темноокрашенных форм бабочки берёзовая пяденица, незаметных для птиц на загрязнённых копотью стволах деревьев (до сер. 19 в. встречалась только светлая форма, имитировавшая пятна лишайников на светлых стволах берёзы). Быстрое привыкание к ядам разл. видов насекомых и грызунов, возникновение резистентности микроорганизмов к антибиотикам свидетельствуют о том, что давления движущего отбора в природных популяциях достаточно для того, чтобы обеспечить быстрый адаптивный ответ на резкие изменения среды. Как правило, селекция по одному признаку влечёт за собой целый ряд преобразований. Напр., длительный отбор на содержание белка или масла в зёрнах кукурузы сопровождается изменениями форм зёрен, размеров початков, их расположения над уровнем почвы и др.
Классич. примером его действия стало преобладание в пром. районах темноокрашенных форм бабочки берёзовая пяденица, незаметных для птиц на загрязнённых копотью стволах деревьев (до сер. 19 в. встречалась только светлая форма, имитировавшая пятна лишайников на светлых стволах берёзы). Быстрое привыкание к ядам разл. видов насекомых и грызунов, возникновение резистентности микроорганизмов к антибиотикам свидетельствуют о том, что давления движущего отбора в природных популяциях достаточно для того, чтобы обеспечить быстрый адаптивный ответ на резкие изменения среды. Как правило, селекция по одному признаку влечёт за собой целый ряд преобразований. Напр., длительный отбор на содержание белка или масла в зёрнах кукурузы сопровождается изменениями форм зёрен, размеров початков, их расположения над уровнем почвы и др.
Результатом действия движущего отбора в филогенезе крупных таксонов является ортоселекция, примером которой служит установленная В.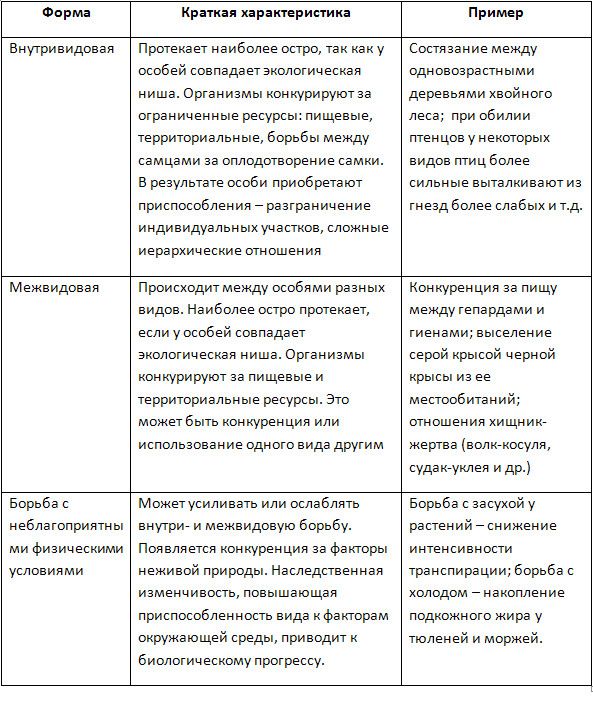 О. Ковалевским направленная эволюция конечности предков лошади (от пятипалости к однопалости), протекавшая миллионы лет и обеспечившая увеличение скорости и экономичности бега.
О. Ковалевским направленная эволюция конечности предков лошади (от пятипалости к однопалости), протекавшая миллионы лет и обеспечившая увеличение скорости и экономичности бега.
Дизруптивный, или разрывающий, отбор благоприятствует сохранению крайних отклонений и ведёт к увеличению полиморфизма. Он проявляется в тех случаях, когда ни одна из внутривидовых форм с разными генотипами не получает абсолютного преимущества в борьбе за существование из-за разнообразия условий, одновременно встречающихся на одной территории; при этом прежде всего элиминируются особи со средним или промежуточным характером признаков. Ещё в нач. 20 в. рос. ботаник Н. В. Цингер показал, что большой погремок (Alectoroleophus major), цветущий и плодоносящий на некошеных лугах в течение всего лета, на скашиваемых лугах образует две расы: ранневесеннюю, успевающую принести семена до начала покоса, и позднеосеннюю – низкие растения, не повреждаемые при покосе, а затем быстро зацветающие и успевающие дать семена до начала морозов. Другим примером полиморфизма служит различие в окраске раковин у земляной улитки (Capaсea nemoralis), являющейся кормом для птиц: в густых буковых лесах, где в течение всего года сохраняется подстилка из красно-бурого опада, обычны особи с коричневой и розовой окраской; на лугах с жёлтой подстилкой преобладают улитки с жёлтой окраской. В смешанных же лиственных лесах, где характер фона меняется с наступлением нового сезона, ранней весной доминируют улитки с бурой и розовой окраской, а летом – с жёлтой. Дарвиновы вьюрки (Geospizinae) на островах Галапагос (классич. пример адаптивной радиации) – конечный результат длительного дизруптивного отбора, приведшего к образованию десятков близкородственных видов.
Другим примером полиморфизма служит различие в окраске раковин у земляной улитки (Capaсea nemoralis), являющейся кормом для птиц: в густых буковых лесах, где в течение всего года сохраняется подстилка из красно-бурого опада, обычны особи с коричневой и розовой окраской; на лугах с жёлтой подстилкой преобладают улитки с жёлтой окраской. В смешанных же лиственных лесах, где характер фона меняется с наступлением нового сезона, ранней весной доминируют улитки с бурой и розовой окраской, а летом – с жёлтой. Дарвиновы вьюрки (Geospizinae) на островах Галапагос (классич. пример адаптивной радиации) – конечный результат длительного дизруптивного отбора, приведшего к образованию десятков близкородственных видов.
Если указанные формы Е. о. приводят к изменению и фенотипич., и генетич. структуры популяций, то впервые описанный И. И. Шмальгаузеном (1938) стабилизирующий отбор сохраняет в популяции среднее значение признаков (норму) и не пропускает в следующее поколение геномы особей, наиболее отклоняющихся от этой нормы. Он направлен на поддержание и повышение устойчивости в популяции среднего, ранее сложившегося фенотипа. Известно, напр., что во время снежных бурь выживают птицы, которые по мн. признакам (длина крыла, клюва, масса тела и т. д.) приближаются к средней норме, а отклонившиеся от этой нормы особи погибают. Размеры и форма цветков у растений, опыляемых насекомыми, более устойчивы, чем у растений, опыляемых ветром, что обусловлено сопряжённой эволюцией растений и их опылителей, «выбраковкой» уклонившихся от нормы форм (напр., шмель не может проникнуть в слишком узкий венчик цветка, а хоботок бабочки не касается слишком коротких тычинок у растений с длинным венчиком).
Шмальгаузеном (1938) стабилизирующий отбор сохраняет в популяции среднее значение признаков (норму) и не пропускает в следующее поколение геномы особей, наиболее отклоняющихся от этой нормы. Он направлен на поддержание и повышение устойчивости в популяции среднего, ранее сложившегося фенотипа. Известно, напр., что во время снежных бурь выживают птицы, которые по мн. признакам (длина крыла, клюва, масса тела и т. д.) приближаются к средней норме, а отклонившиеся от этой нормы особи погибают. Размеры и форма цветков у растений, опыляемых насекомыми, более устойчивы, чем у растений, опыляемых ветром, что обусловлено сопряжённой эволюцией растений и их опылителей, «выбраковкой» уклонившихся от нормы форм (напр., шмель не может проникнуть в слишком узкий венчик цветка, а хоботок бабочки не касается слишком коротких тычинок у растений с длинным венчиком).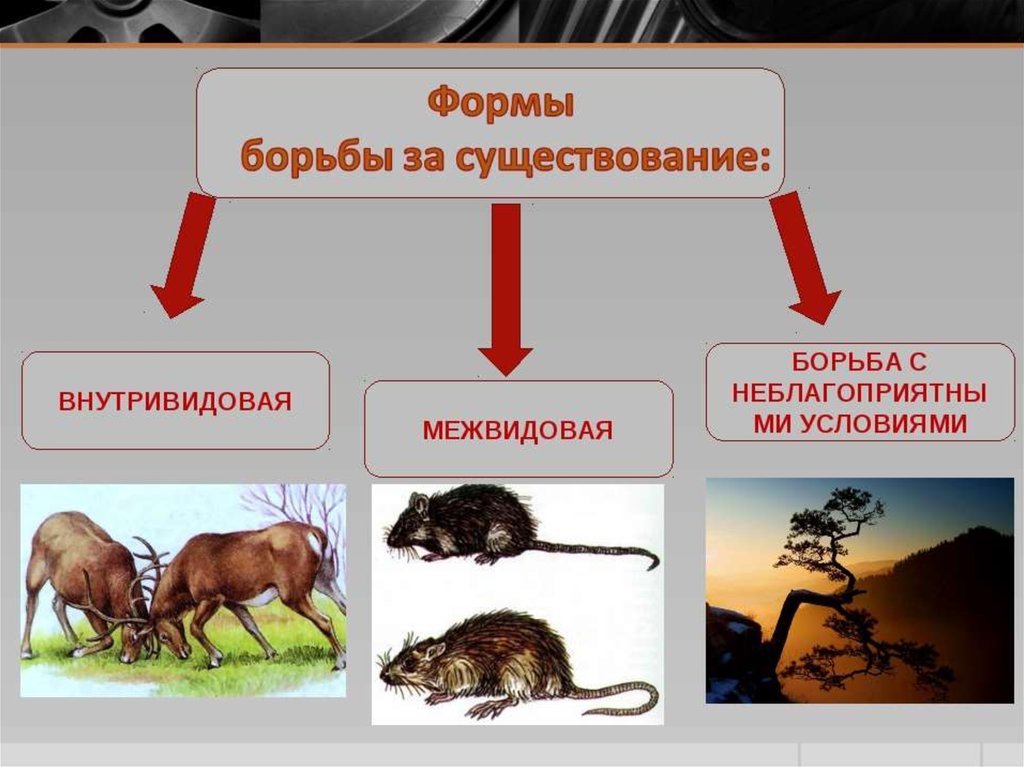 Благодаря стабилизирующему отбору при внешнем неизменном фенотипе могут идти существенные генетич. изменения, обеспечивающие независимость развития адаптаций от колеблющихся условий среды. Одним из результатов действия стабилизирующего отбора можно считать «биохимическую универсальность» жизни на Земле.
Благодаря стабилизирующему отбору при внешнем неизменном фенотипе могут идти существенные генетич. изменения, обеспечивающие независимость развития адаптаций от колеблющихся условий среды. Одним из результатов действия стабилизирующего отбора можно считать «биохимическую универсальность» жизни на Земле.
Дестабилизирующий отбор (назв. предложено Д. К. Беляевым, 1970) ведёт к резкому нарушению систем регуляции онтогенеза, вскрытию мобилизационного резерва и росту фенотипич. изменчивости при интенсивном отборе в каком-либо определённом направлении. Напр., отбор на снижение агрессивности хищных зверей в неволе через перестройку нейрогуморальной системы ведёт к дестабилизации цикла размножения, сдвигам в сроках линьки, изменениям в положении хвоста, ушей, в окраске и др.
Обнаружены гены, которые могут быть летальными или снижать жизнеспособность организмов в гомозиготном состоянии, а в гетерозиготном, напротив, повышать экологич.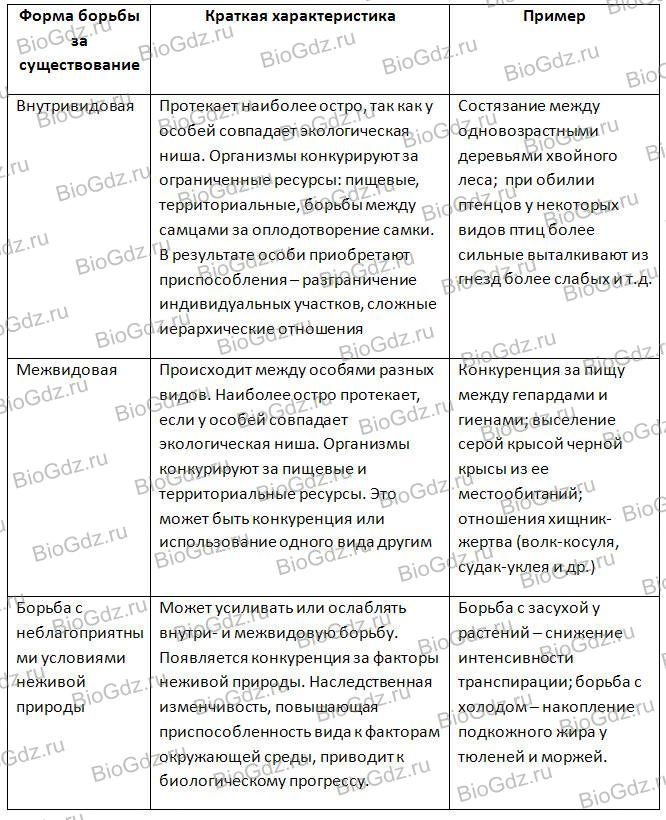 пластичность и др. показатели. В этом случае можно говорить о т. н. сбалансированном отборе, обеспечивающем поддержание генетич. разнообразия с определённым соотношением частот аллелей. Примером его действия может служить повышение устойчивости у больных серповидно-клеточной анемией (гетерозиготных по гену гемоглобина S) к заражению разл. штаммами малярийного плазмодия (см. Гемоглобины).
пластичность и др. показатели. В этом случае можно говорить о т. н. сбалансированном отборе, обеспечивающем поддержание генетич. разнообразия с определённым соотношением частот аллелей. Примером его действия может служить повышение устойчивости у больных серповидно-клеточной анемией (гетерозиготных по гену гемоглобина S) к заражению разл. штаммами малярийного плазмодия (см. Гемоглобины).
Важным шагом в преодолении стремления все признаки организмов объяснять действием Е. о. стала концепция нейтральной эволюции, согласно которой часть изменений на уровне белков и нуклеиновых кислот происходит путём фиксации адаптивно нейтральных или почти нейтральных мутаций. Возможен отбор видов, возникающих в периферийных популяциях «внезапно» с геохронологич. точки зрения. Ещё раньше было доказано, что катастрофический отбор, при котором в период резких изменений среды выживает небольшое число особей и даже единственный организм, может стать основой формирования нового вида за счёт хромосомной перестройки и смены экологич. ниши. Так, образование ксерофитного, эндемичного вида Clarkia lingulata в горах Сьерра-Невада в Калифорнии объясняют сильной засухой, вызвавшей массовую гибель растений, принявшую катастрофич. характер в периферийных популяциях.
ниши. Так, образование ксерофитного, эндемичного вида Clarkia lingulata в горах Сьерра-Невада в Калифорнии объясняют сильной засухой, вызвавшей массовую гибель растений, принявшую катастрофич. характер в периферийных популяциях.
Е. о., затрагивающий вторичные половые признаки особей, называют половым (напр., яркая брачная окраска самцов у мн. видов рыб и птиц, зазывающие крики, специфич. запахи, сильно развитые орудия для турнирного боя у млекопитающих). Эти признаки полезны, т. к. повышают возможность участия их носителей в воспроизведении потомства. В половом отборе наибольшую активность проявляют самцы, что выгодно для вида в целом, т. к. самки остаются в большей безопасности в период размножения.
Выделяют также групповой отбор, способствующий сохранению признаков, полезных семье, стае, колонии. Его частным случаем у колониальных насекомых является отбор сородичей, при котором стерильные касты (рабочие, солдаты и др. ) обеспечивают (нередко ценой собственной жизни) выживание плодовитых особей (маток) и личинок и тем самым сохранение всей колонии. Альтруистическое поведение родителей, притворяющихся ранеными, чтобы увести хищника от своих детей, грозит гибелью имитатору, но в целом повышает шансы на выживание его потомства.
) обеспечивают (нередко ценой собственной жизни) выживание плодовитых особей (маток) и личинок и тем самым сохранение всей колонии. Альтруистическое поведение родителей, притворяющихся ранеными, чтобы увести хищника от своих детей, грозит гибелью имитатору, но в целом повышает шансы на выживание его потомства.
Хотя представления о ведущей роли Е. о. в эволюции получили подтверждение во множестве экспериментов, до сих пор они подвергаются критике, исходящей из представления о невозможности образования организмов в результате случайной комбинации мутаций. При этом игнорируется тот факт, что каждый акт Е. о. совершается на базе предыдущих результатов его же действия, которые, в свою очередь, предопределяют формы, интенсивность и направления Е. о., а значит, пути и закономерности эволюции.
О происхождении видов: Глава III, Борьба за существование | Чарльз Дарвин
Дарвин уже показал своему читателю силу искусственного отбора и изощренные вариации, существующие в природе. В главе III он спрашивает, как разновидности могут стать полноценными видами. Решающее значение для этого имеет то, что он называет «борьбой за существование». Это может быть конфликт из-за еды или борьба со стихией. Одним из доказательств этой борьбы является огромный потенциал роста популяций животных и растений. Даже слон, один из самых медленно размножающихся в животном мире, мог бы захватить планету, если бы ему было позволено беспрепятственно размножаться в течение достаточно долгого времени. По словам Дарвина, через 500 лет одна пара оставит 15 миллионов потомков. Популяции контролируются, потому что не хватает ресурсов, и это приводит к острой конкуренции за выживание.
В главе III он спрашивает, как разновидности могут стать полноценными видами. Решающее значение для этого имеет то, что он называет «борьбой за существование». Это может быть конфликт из-за еды или борьба со стихией. Одним из доказательств этой борьбы является огромный потенциал роста популяций животных и растений. Даже слон, один из самых медленно размножающихся в животном мире, мог бы захватить планету, если бы ему было позволено беспрепятственно размножаться в течение достаточно долгого времени. По словам Дарвина, через 500 лет одна пара оставит 15 миллионов потомков. Популяции контролируются, потому что не хватает ресурсов, и это приводит к острой конкуренции за выживание.
Прежде чем перейти к предмету этой главы, я должен сделать несколько предварительных замечаний, чтобы показать, как борьба за существование связана с естественным отбором. В предыдущей главе было замечено, что среди органических существ в естественном состоянии существует некоторая индивидуальная изменчивость; действительно, я не знаю, чтобы это когда-либо оспаривалось. Для нас безразлично, называть ли множество сомнительных форм видами, подвидами или разновидностями; какое место, например, могут занимать две или три сотни сомнительных форм британских растений, если допустить существование каких-либо хорошо заметных разновидностей. Но простое существование индивидуальной изменчивости и нескольких хорошо заметных разновидностей, хотя и необходимое в качестве основы для работы, мало помогает нам понять, как виды возникают в природе. Каким образом были совершены все эти изысканные приспособления одной части организации к другой части и к условиям жизни, а также одного отдельного органического существа к другому существу? Наиболее ясно мы видим эти прекрасные коадаптации у дятла и омелы; и лишь немногим менее очевидно у самого скромного паразита, который цепляется за шерсть четвероногого или за перья птицы; в строении жука, ныряющего под воду; в семени с перьями, которое развевается самым нежным ветерком; Короче говоря, мы видим прекрасные приспособления повсюду и в каждой части органического мира.
Для нас безразлично, называть ли множество сомнительных форм видами, подвидами или разновидностями; какое место, например, могут занимать две или три сотни сомнительных форм британских растений, если допустить существование каких-либо хорошо заметных разновидностей. Но простое существование индивидуальной изменчивости и нескольких хорошо заметных разновидностей, хотя и необходимое в качестве основы для работы, мало помогает нам понять, как виды возникают в природе. Каким образом были совершены все эти изысканные приспособления одной части организации к другой части и к условиям жизни, а также одного отдельного органического существа к другому существу? Наиболее ясно мы видим эти прекрасные коадаптации у дятла и омелы; и лишь немногим менее очевидно у самого скромного паразита, который цепляется за шерсть четвероногого или за перья птицы; в строении жука, ныряющего под воду; в семени с перьями, которое развевается самым нежным ветерком; Короче говоря, мы видим прекрасные приспособления повсюду и в каждой части органического мира.
Опять же могут спросить, как получается, что разновидности, которые я назвал зарождающимися видами, в конце концов превращаются в хорошие и отличные виды, которые в большинстве случаев явно отличаются друг от друга гораздо больше, чем разновидности одного и того же вида? ? Как возникают те группы видов, которые составляют так называемые отдельные роды и отличаются друг от друга больше, чем виды одного и того же рода? Все эти результаты, как мы более полно увидим в следующей главе, неизбежно вытекают из борьбы за жизнь. Благодаря этой борьбе за жизнь всякое изменение, как бы оно ни было незначительно и по какой бы причине оно ни происходило, если оно в какой-либо степени полезно для индивидуума любого вида в его бесконечно сложных отношениях с другими органическими существами и с внешней природой, будет иметь тенденцию к сохранение этого индивидуума и обычно передается по наследству его потомству. Потомство, таким образом, также будет иметь больше шансов на выживание, поскольку из многих особей любого вида, которые периодически рождаются, выжить может лишь небольшое число. Я назвал этот принцип, благодаря которому каждое незначительное изменение, если оно полезно, сохраняется, термином естественного отбора, чтобы обозначить его отношение к способности человека к отбору. Мы видели, что человек посредством отбора, несомненно, может производить большие результаты и может приспособить органические существа к своим собственным потребностям путем накопления незначительных, но полезных вариаций, данных ему рукой Природы. Но естественный отбор, как мы увидим далее, есть сила, постоянно готовая к действию, и она так же неизмеримо превосходит слабые усилия человека, как творения природы превосходят произведения искусства.
Я назвал этот принцип, благодаря которому каждое незначительное изменение, если оно полезно, сохраняется, термином естественного отбора, чтобы обозначить его отношение к способности человека к отбору. Мы видели, что человек посредством отбора, несомненно, может производить большие результаты и может приспособить органические существа к своим собственным потребностям путем накопления незначительных, но полезных вариаций, данных ему рукой Природы. Но естественный отбор, как мы увидим далее, есть сила, постоянно готовая к действию, и она так же неизмеримо превосходит слабые усилия человека, как творения природы превосходят произведения искусства.
………………………………………………………. …………………………………………. ………………………………………….
Теперь мы более подробно обсудим борьбу за существование. В моей будущей работе эта тема будет рассмотрена, как она и заслуживает, гораздо подробнее. Старшие де Кандоль и Лайель широко и философски показали, что все органические существа подвергаются жесткой конкуренции. Что касается растений, никто не занимался этим вопросом с большим энтузиазмом и умением, чем У. Герберт, декан Манчестера, что, очевидно, является результатом его больших знаний в области садоводства. Нет ничего легче, как признать на словах истину всеобщей борьбы за жизнь, и нет ничего труднее, — по крайней мере, я нашел это таковым, — чем постоянно иметь в виду этот вывод. Тем не менее, если это не будет полностью укоренено в уме, я убежден, что вся экономия природы, с каждым фактом о распространении, редкости, изобилии, исчезновении и изменчивости, будет видна смутно или совершенно неправильно понята. Мы созерцаем лицо природы, сияющее радостью, мы часто видим изобилие пищи; мы не видим или забываем, что птицы, праздно поющие вокруг нас, питаются большей частью насекомыми или семенами и таким образом постоянно губят жизнь; или мы забываем, как сильно эти певчие, или их яйца, или их птенцы уничтожаются птицами и хищными зверями; мы не всегда помним, что, хотя сейчас пища может быть в изобилии, это не так во все времена года каждого повторяющегося года.
Что касается растений, никто не занимался этим вопросом с большим энтузиазмом и умением, чем У. Герберт, декан Манчестера, что, очевидно, является результатом его больших знаний в области садоводства. Нет ничего легче, как признать на словах истину всеобщей борьбы за жизнь, и нет ничего труднее, — по крайней мере, я нашел это таковым, — чем постоянно иметь в виду этот вывод. Тем не менее, если это не будет полностью укоренено в уме, я убежден, что вся экономия природы, с каждым фактом о распространении, редкости, изобилии, исчезновении и изменчивости, будет видна смутно или совершенно неправильно понята. Мы созерцаем лицо природы, сияющее радостью, мы часто видим изобилие пищи; мы не видим или забываем, что птицы, праздно поющие вокруг нас, питаются большей частью насекомыми или семенами и таким образом постоянно губят жизнь; или мы забываем, как сильно эти певчие, или их яйца, или их птенцы уничтожаются птицами и хищными зверями; мы не всегда помним, что, хотя сейчас пища может быть в изобилии, это не так во все времена года каждого повторяющегося года.
Должен предположить, что я использую термин Борьба за Существование в широком и метафорическом смысле, включая зависимость одного существа от другого и включая (что более важно) не только жизнь индивидуума, но и успех в оставлении потомства. Можно сказать, что два псовых во время голода борются друг с другом, чтобы получить пищу и жить. Но говорят, что растение на краю пустыни борется за жизнь с засухой, хотя правильнее было бы сказать, что оно зависит от влаги. О растении, которое ежегодно производит тысячу семян, из которых в среднем только одно достигает зрелости, можно с большей уверенностью сказать, что оно борется с растениями того же и других видов, уже покрывающими землю. Омела зависит от яблони и некоторых других деревьев, но можно сказать, что она борется с этими деревьями лишь в надуманном смысле, потому что, если на одном дереве вырастет слишком много этих паразитов, оно зачахнет и погибнет. Но несколько сеянцев омелы, растущих близко друг к другу на одной и той же ветке, правильнее было бы сказать, что они борются друг с другом. Поскольку омелу распространяют птицы, ее существование зависит от птиц; метафорически можно сказать, что оно борется с другими плодоносящими растениями, чтобы соблазнить птиц пожрать и таким образом распространить его семена, а не семена других растений. В этих нескольких смыслах, переходящих друг в друга, я использую для удобства общий термин борьбы за существование.
Поскольку омелу распространяют птицы, ее существование зависит от птиц; метафорически можно сказать, что оно борется с другими плодоносящими растениями, чтобы соблазнить птиц пожрать и таким образом распространить его семена, а не семена других растений. В этих нескольких смыслах, переходящих друг в друга, я использую для удобства общий термин борьбы за существование.
Борьба за существование неизбежно вытекает из высокой скорости, с которой имеют тенденцию размножаться все органические существа. Каждое существо, которое в течение своей естественной жизни производит несколько яиц или семян, должно подвергаться уничтожению в какой-то период своей жизни, в какое-то время года или в какой-то случайный год, иначе, по принципу геометрического увеличения, его численность быстро стала бы настолько непомерно велика, что ни одна страна не может поддерживать продукт. Следовательно, так как производится больше особей, чем может выжить, то во всяком случае должна иметь место борьба за существование либо одной особи с другой, принадлежащей к тому же виду, либо с особями различных видов, либо с физическими условиями жизни. Это учение Мальтуса, примененное с многократным усилием ко всему животному и растительному царствам; ибо в этом случае не может быть ни искусственного увеличения пищи, ни благоразумного воздержания от брака. Хотя в настоящее время численность некоторых видов может увеличиваться более или менее быстро, все не могут этого сделать, ибо мир не удержит их.
Это учение Мальтуса, примененное с многократным усилием ко всему животному и растительному царствам; ибо в этом случае не может быть ни искусственного увеличения пищи, ни благоразумного воздержания от брака. Хотя в настоящее время численность некоторых видов может увеличиваться более или менее быстро, все не могут этого сделать, ибо мир не удержит их.
………………………………………………………. …………………………………………. ………………………………………….
Нет никаких исключений из правила, согласно которому каждое органическое существо естественным образом размножается с такой высокой скоростью, что, если бы оно не было уничтожено, земля вскоре была бы покрыта потомством одной пары. Даже медленно размножающийся человек удвоился за 25 лет, и при таких темпах через несколько тысяч лет у его потомства буквально не осталось бы места. Линней подсчитал, что если бы однолетнее растение дало только два семени — а нет растения столь непродуктивного, как это, — а их сеянцы в следующем году дали бы два и т. д., то через двадцать лет было бы миллион растений. Слон считается самым медленным размножающимся из всех известных животных, и я приложил некоторые усилия, чтобы оценить вероятную минимальную норму его естественного прироста: было бы недобросовестно предположить, что он размножается в возрасте тридцати лет и продолжает размножаться. до девяноста лет, принося за этот промежуток три пары детенышей; если бы это было так, то в конце пятого века было бы живо пятнадцать миллионов слонов, потомков первой пары.
д., то через двадцать лет было бы миллион растений. Слон считается самым медленным размножающимся из всех известных животных, и я приложил некоторые усилия, чтобы оценить вероятную минимальную норму его естественного прироста: было бы недобросовестно предположить, что он размножается в возрасте тридцати лет и продолжает размножаться. до девяноста лет, принося за этот промежуток три пары детенышей; если бы это было так, то в конце пятого века было бы живо пятнадцать миллионов слонов, потомков первой пары.
Но у нас есть лучшие доказательства по этому вопросу, чем чисто теоретические расчеты, а именно многочисленные зарегистрированные случаи поразительно быстрого роста различных животных в естественном состоянии, когда обстоятельства были благоприятны для них в течение двух или трех последующих сезонов. Еще более поразительны данные, полученные от многих видов наших домашних животных, которые одичали в некоторых частях света: если бы заявления о темпах роста медленно размножающегося крупного рогатого скота и лошадей в Южной Америке, а в последнее время и в Австралии, не были бы хорошо аутентифицированы, они были бы совершенно невероятными.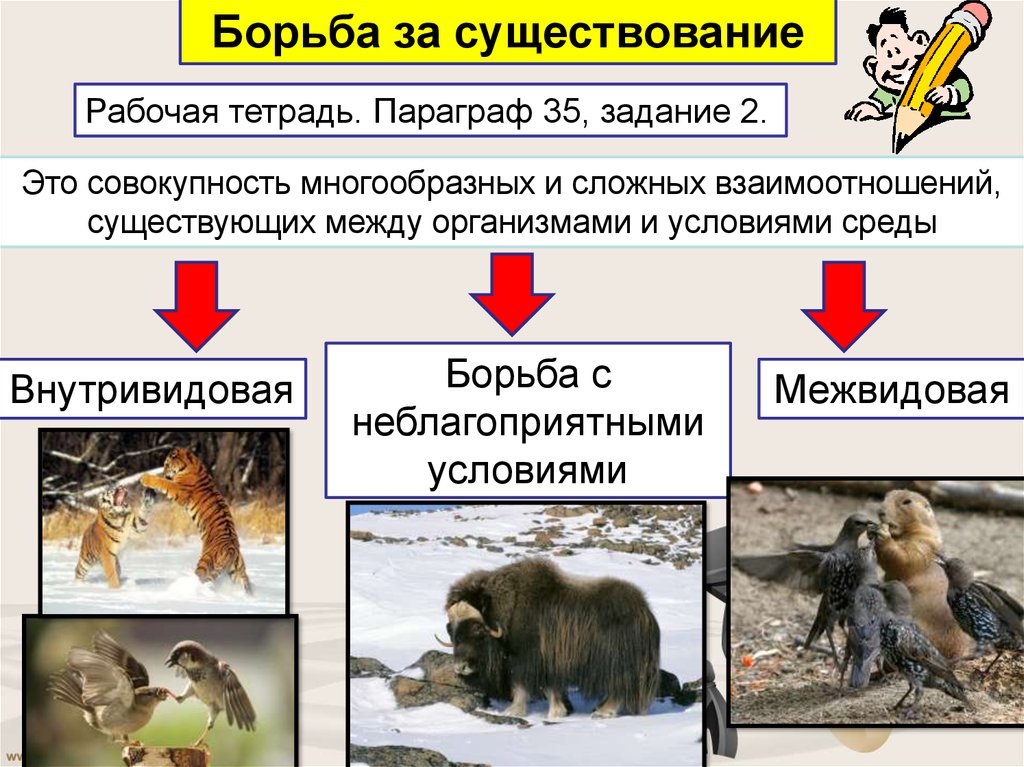 То же самое и с растениями: можно привести случаи интродуцированных растений, которые стали обычными на целых островах менее чем за десять лет. Некоторые из растений, наиболее многочисленных в настоящее время на широких равнинах Ла-Платы, покрывающих квадратные лиги поверхности почти за исключением всех других растений, были завезены из Европы; и есть растения, которые в настоящее время распространены в Индии, как я слышал от доктора Фальконера, от мыса Коморин до Гималаев, которые были импортированы из Америки с момента ее открытия. В таких случаях, а примеров можно было бы привести бесконечное количество, никто не предполагает, что плодовитость этих животных или растений внезапно и временно увеличилась в какой-либо заметной степени. Очевидное объяснение состоит в том, что жизненные условия были очень благоприятны, и, следовательно, было меньше истребления старых и молодых, и что почти все молодые были способны к размножению. В таких случаях геометрическая пропорция увеличения, результат которого всегда удивляет, просто объясняет необычайно быстрое увеличение и широкое распространение натурализованных продуктов в их новых домах.
То же самое и с растениями: можно привести случаи интродуцированных растений, которые стали обычными на целых островах менее чем за десять лет. Некоторые из растений, наиболее многочисленных в настоящее время на широких равнинах Ла-Платы, покрывающих квадратные лиги поверхности почти за исключением всех других растений, были завезены из Европы; и есть растения, которые в настоящее время распространены в Индии, как я слышал от доктора Фальконера, от мыса Коморин до Гималаев, которые были импортированы из Америки с момента ее открытия. В таких случаях, а примеров можно было бы привести бесконечное количество, никто не предполагает, что плодовитость этих животных или растений внезапно и временно увеличилась в какой-либо заметной степени. Очевидное объяснение состоит в том, что жизненные условия были очень благоприятны, и, следовательно, было меньше истребления старых и молодых, и что почти все молодые были способны к размножению. В таких случаях геометрическая пропорция увеличения, результат которого всегда удивляет, просто объясняет необычайно быстрое увеличение и широкое распространение натурализованных продуктов в их новых домах.
Посмотрите на растение посреди своего ареала, почему оно не удваивает и не учетверяет свои номера? Мы знаем, что он вполне может выдерживать немного больше жары или холода, сырости или сухости, ибо в других местах он колеблется в более жарких или холодных, влажных или более сухих районах. В этом случае мы можем ясно видеть, что если бы мы хотели в воображении дать растению способность увеличиваться в числе, мы должны были бы дать ему некоторое преимущество перед его конкурентами или над животными, которые охотятся на него. В пределах своего географического ареала изменение конституции по отношению к климату явно было бы преимуществом для нашего растения; но у нас есть основания полагать, что лишь немногие растения или животные обитают так далеко, что они погибают от одной лишь суровости климата. Конкуренция прекратится только тогда, когда мы достигнем крайних пределов жизни, арктических регионов или границ абсолютной пустыни. Земля может быть очень холодной или сухой, но между несколькими видами или между особями одного и того же вида будет конкуренция за самые теплые или самые влажные места.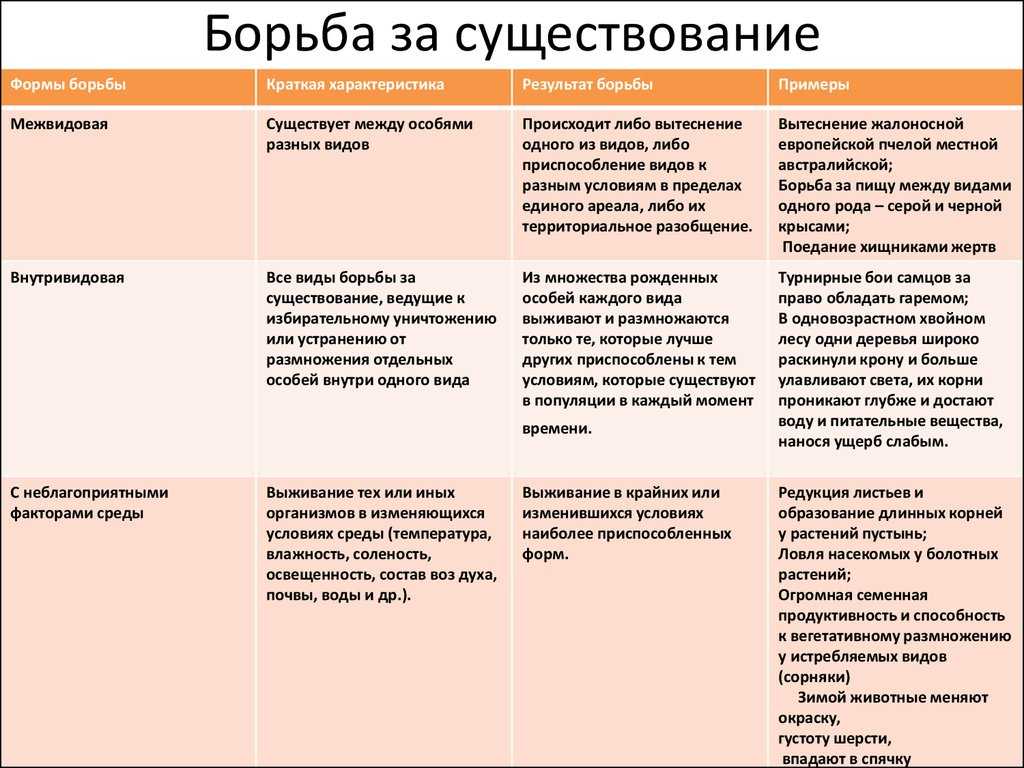
………………………………………………………. …………………………………………. ………………………………………….
Отсюда мы также можем видеть, что когда растение или животное помещается в новую страну среди новых конкурентов, хотя климат может быть точно таким же, как на его прежней родине, но условия его жизни обычно изменяются в существенной манере. Если бы мы хотели увеличить его среднюю численность в его новом доме, мы должны были бы изменить его способом, отличным от того, что мы должны были бы сделать в его родной стране; ибо мы должны были бы дать ему какое-то преимущество перед другим набором конкурентов или врагов.
Таким образом, хорошо попытаться в своем воображении придать какой-либо форме некоторое преимущество перед другой. Вероятно, ни в одном отдельном случае мы не должны знать, что делать, чтобы добиться успеха. Это убедит нас в нашем невежестве во взаимоотношениях всех органических существ; судимость настолько необходима, насколько трудно ее приобрести.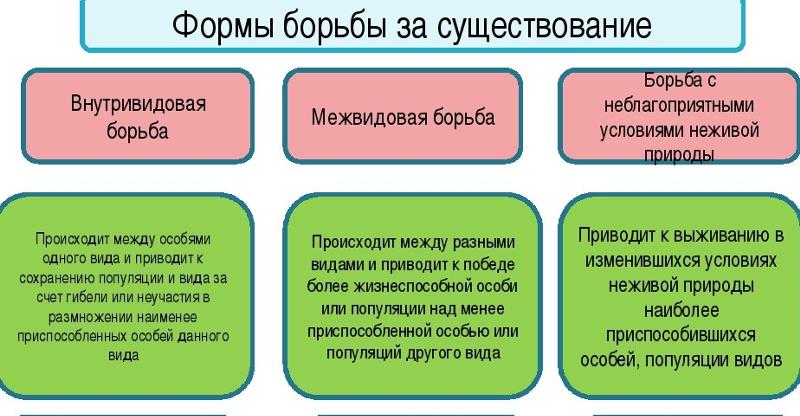 Все, что мы можем сделать, это постоянно помнить, что каждое органическое существо стремится увеличиться в геометрической прогрессии; что каждому в какой-то период своей жизни, в какое-то время года, в каждом поколении или через определенные промежутки времени приходится бороться за жизнь и терпеть великие разрушения. Когда мы размышляем об этой борьбе, мы можем утешаться полной верой в то, что война природы не непрестанна, что не чувствуется страха, что смерть обычно близка и что сильные, здоровые и счастливые выживают и умножить.
Все, что мы можем сделать, это постоянно помнить, что каждое органическое существо стремится увеличиться в геометрической прогрессии; что каждому в какой-то период своей жизни, в какое-то время года, в каждом поколении или через определенные промежутки времени приходится бороться за жизнь и терпеть великие разрушения. Когда мы размышляем об этой борьбе, мы можем утешаться полной верой в то, что война природы не непрестанна, что не чувствуется страха, что смерть обычно близка и что сильные, здоровые и счастливые выживают и умножить.
Прочтите следующий отрывок: Глава IV, Естественный отбор
 Дарвин утверждает, что каждый вид будет расти с экспоненциальной скоростью, но ограниченное количество имеющихся ресурсов создаст постоянную борьбу за существование между представителями вида, поскольку каждый индивидуум конкурирует за получение необходимых ресурсов и производство потомства. Как более
производятся особи, чем они могут выжить, должно быть
каждый случай будет борьбой за существование, либо один человек с
другой того же вида, или с особями различных
вида или с физическими условиями жизни. В результате во всей природе происходит большое уничтожение жизни, поскольку не все организмы преуспевают в борьбе за существование. Учитывая, что Дарвин устанавливает в главе 2, что существуют некоторые вариации в природе, он сочетает эту идею с наличием борьбы за существование, чтобы выдвинуть идею о том, что определенные вариации в организмах дадут преимущества этому организму и позволят этому индивиду добиться успеха. в борьбе за существование и пережить своих соперников. Этот индивидуум произведет потомство и, таким образом, передаст эти благоприятные вариации, в то время как в то же время те индивидуумы без таких вариаций вымрут и не смогут передать свою собственную ДНК.
Дарвин утверждает, что каждый вид будет расти с экспоненциальной скоростью, но ограниченное количество имеющихся ресурсов создаст постоянную борьбу за существование между представителями вида, поскольку каждый индивидуум конкурирует за получение необходимых ресурсов и производство потомства. Как более
производятся особи, чем они могут выжить, должно быть
каждый случай будет борьбой за существование, либо один человек с
другой того же вида, или с особями различных
вида или с физическими условиями жизни. В результате во всей природе происходит большое уничтожение жизни, поскольку не все организмы преуспевают в борьбе за существование. Учитывая, что Дарвин устанавливает в главе 2, что существуют некоторые вариации в природе, он сочетает эту идею с наличием борьбы за существование, чтобы выдвинуть идею о том, что определенные вариации в организмах дадут преимущества этому организму и позволят этому индивиду добиться успеха. в борьбе за существование и пережить своих соперников. Этот индивидуум произведет потомство и, таким образом, передаст эти благоприятные вариации, в то время как в то же время те индивидуумы без таких вариаций вымрут и не смогут передать свою собственную ДНК.
 При постоянном увеличении популяций всех видов такая крайняя гибель людей имеет решающее значение, иначе мир был бы покрыт и во власти, как предполагает Дарвин, потомства всего одной пары. Хотя этот тяжелый фактор смерти может показаться суровым и печальным, Дарвин напоминает читателю, что «мы можем утешаться полной верой в то, что война природы не непрестанна, что не чувствуется страха, что смерть обычно немедленна и что сильные, здоровые и счастливые выживают и размножаются».
При постоянном увеличении популяций всех видов такая крайняя гибель людей имеет решающее значение, иначе мир был бы покрыт и во власти, как предполагает Дарвин, потомства всего одной пары. Хотя этот тяжелый фактор смерти может показаться суровым и печальным, Дарвин напоминает читателю, что «мы можем утешаться полной верой в то, что война природы не непрестанна, что не чувствуется страха, что смерть обычно немедленна и что сильные, здоровые и счастливые выживают и размножаются».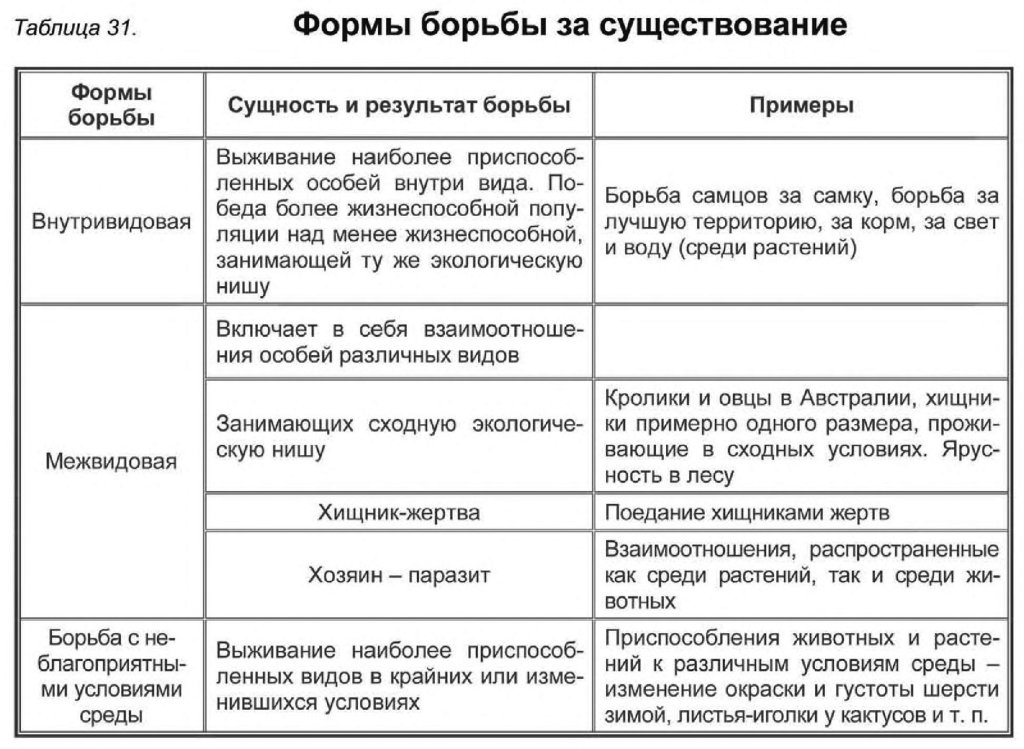 Вместо этого большое количество яиц или семян компенсирует большее количество разрушений в более позднем возрасте, обычно молодых. Если погибает много семян или молодняка, то их должно быть произведено много, чтобы вид оставался неповрежденным и стабильным. Дарвин цитирует: «достаточно было бы поддерживать полную численность дерева, живущего в среднем тысячу лет, если бы одно семя производилось раз в тысячу лет, если предположить, что это семя никогда не уничтожалось». Таким образом, количество потомков данного вида напрямую не влияет на среднюю численность этого вида.
Вместо этого большое количество яиц или семян компенсирует большее количество разрушений в более позднем возрасте, обычно молодых. Если погибает много семян или молодняка, то их должно быть произведено много, чтобы вид оставался неповрежденным и стабильным. Дарвин цитирует: «достаточно было бы поддерживать полную численность дерева, живущего в среднем тысячу лет, если бы одно семя производилось раз в тысячу лет, если предположить, что это семя никогда не уничтожалось». Таким образом, количество потомков данного вида напрямую не влияет на среднюю численность этого вида.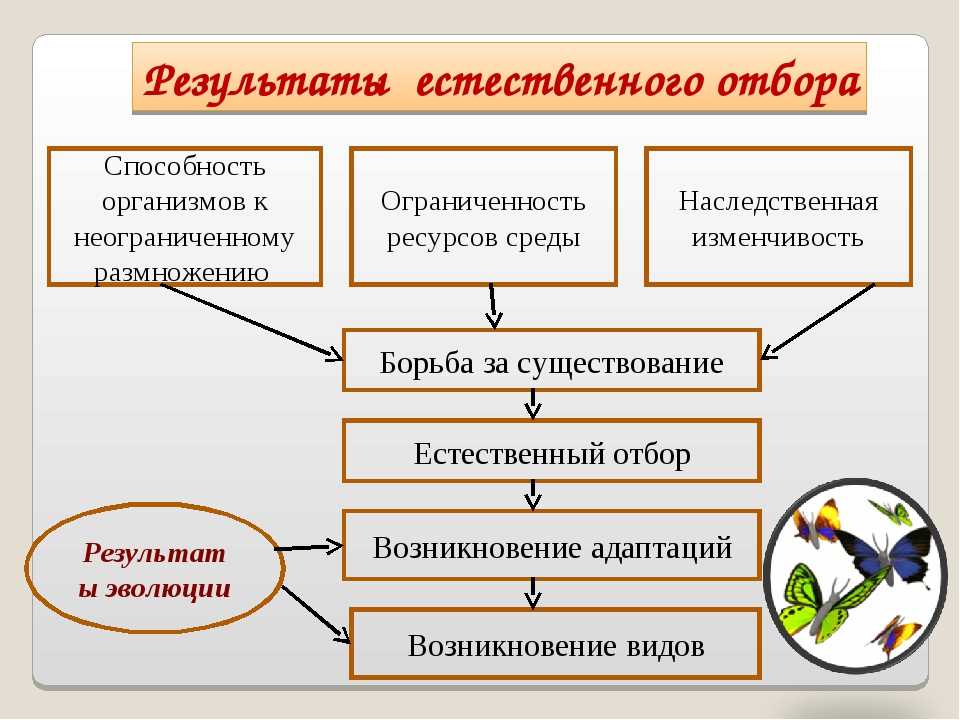
Leave A Comment