
d09ad0bed180d0b5d0bdd18c
Share to:
Summary «%d0%9a%d0%be%d1%80%d0%b5%d0%bd%d1%8c»: sman 1 cisarua sma parulian 1 medan smp 1 jepon sd negeri tamansari 1 yogyakarta smp negeri 2 rajeg smp negeri 2 kota cirebon alwi smith rt rw g. obos 12 logo smk yadika manado fransiskus xaverius teguh smp negeri 242 jakarta foto smp muhammadiyah 1 wates gkps tanjung morawa logo sman 1 binuang meguri logo sma n 1 maos billa film sma 6 wajo sma negeri 53 jakarta foto stanag 4569 super smash bros. untuk nintendo 3ds dan wii u sd marhamah pocong sd sdit insan rahayu profil sman 1 malingping sman 1 nalumsari tk nurul iman bandung jaka fajar fatriansyah sd muhammadiyah 1 muntilan ivan dorschner sd…
n tropodo 2 foto peristiwa carrington smp it al fadl maila dinia husni rahiem ka sri tanjung sd marsudirini bsb semarang another one bites the dust tarombo marga simanjuntak oliver wendell holmes rokok lucky strike cool switch bet365.
ADV (Himnario Adventista)
AG (Aradhana Geethamulu)
AYM (Anointed Youth Missioners)
BE (Buku Ende)
BN (Buku Nyanyian HKBP)
ELI1 (ELI ABOLOJO (Christian Songs, Igala))
ELI2 (ELI KEKE (Short Songs, Igala))
EN (English Hymns)
GB (Gita Bakti)
GBL (Gita Bakti Edisi Lama)
HC (Держись Христа)
JB (Jiwaku Bersukacita: Pujian Sekolah Minggu)
KJ (Kidung Jemaat)
KLIK (Kumpulan Lagu Ibadah Kreatif dan KKR)
KPJ (Kidung Pasamuan Jawi)
KPKA (Kidung Pasamuan Kristen Anyar)
KPKL (Kidung Pasamuan Kristen Lawas)
KPPK (Kidung Puji-Pujian Kristen)
KPRI (Kidung Persekutuan Reformed Injili 2004)
KRI (Kidung Reformed Injili 2017 (Tidak resmi))
LS (Lagu Sion Edisi Lengkap)
MNR1 (Mazmur & Nyanyian Rohani Buku 1 (Mazmur))
MNR2 (Mazmur & Nyanyian Rohani Buku 2 (Nyanyian Rohani))
MSDAH (ဧဝံဂေလိဓမ္မသီချင်း (Myanmar Gospel Hymnal))
NKB (Nyanyikanlah Kidung Baru)
NKI (Nyanyian Kemenangan Iman)
NNBT (Nyanyikanlah Nyanyian Baru bagi Tuhan)
NP (Nyanyian Pujian)
NR (Nafiri Rohani)
NRM (Nyanyian Rohani Methodist)
PKJ (Pelengkap Kidung Jemaat)
PKS (Pwuhken Koul Sarawi)
PPK (Puji-pujian Kristen)
PPPR (Puji-pujian Pemuda/Remaja)
RRZ (Runyankole Rukiga, Zaburi)
SP (Песнь Хвалы)
SPSS (Śpiewajmy Panu wyd. dziesiąte)
dziesiąte)
SR (Песнь Возрождения)
SR3300 (Песнь Возрождения 3300 / Song of Revival 3300)
SS (Духовни Песни)
SY (Песни Юности. Том 1 (Songs of Youth, Tome 1))
TSDAH (Worship Songs Hymnal (เพลงสรรเสริญ))
YJ (Юность-Иисусу)
YSMS (Тебе пою, о мой Спаситель)
41
REAKSI vs RESPONS
REAKSI vs RESPONSCEO Google, Sundar Pichai mulai banyak dikenal orang setelah menjabat pimpinan tertinggi raksasa perusa…
CPU (Centra Proccesing Unit)
Pengertian CPUMungkin kita kadang bertanya, bagaimana komputer bisa mengolah data dengan begitu cepat dan tepat? Jawaban…
Pendudukan Jepang di Indonesia
1. Masuknya Jepang Ke IndonesiaTentu, kalian masih ingat bahwa Jepang dengan mudah berhasil menguasai daerah-daera…
5 Obat Mujarab untuk Penyakit Kulit
Obat penyakit kulit harus digunakan sesuai penyebab atau jenis penyakit kulit agar efektif mengatasi keluhan yang terjad…
Berikut hasil pencarian kami kepada anda terkait artikel %d0%9a%d0%be%d1%80%d0%b5%d0%bd%d1%8c :
sman 1 cisarua
sma parulian 1 medan
smp 1 jepon
sd negeri tamansari 1 yogyakarta
smp negeri 2 rajeg
smp negeri 2 kota cirebon
alwi smith
rt rw g.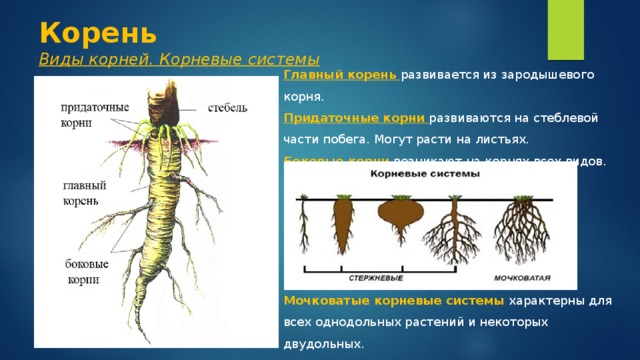 obos 12
obos 12
logo smk yadika manado
fransiskus xaverius teguh
smp negeri 242 jakarta foto
smp muhammadiyah 1 wates
gkps tanjung morawa
logo sman 1 binuang
meguri
logo sma n 1 maos
billa film
sma 6 wajo
sma negeri 53 jakarta foto
stanag 4569
super smash bros. untuk nintendo 3ds dan wii u
sd marhamah
pocong sd
sdit insan rahayu
profil sman 1 malingping
sman 1 nalumsari
tk nurul iman bandung
jaka fajar fatriansyah
sd muhammadiyah 1 muntilan
ivan dorschner
sdn tropodo 2 foto
peristiwa carrington
smp it al fadl
maila dinia husni rahiem
ka sri tanjung
sd marsudirini bsb semarang
another one bites the dust
tarombo marga simanjuntak
oliver wendell holmes
rokok lucky strike cool switch
bet365.com
tkit puzzle
sman 10 padang
umar surya fana
wisma car life insurance
slb jambi
al kahfi islamic school batam
istilah anatomi tulang
smkn 7 yogyakarta
sma bahrul ulum tambakberas jombang
ibrani 8 ayat 12
smp islam ar rahmah jakarta
yoon yoo-sun
gunung waringin
smpn 1 ciawi tasikmalaya
sdn galunggung tasikmalaya
sd negeri pabuaran 01
sd negeri setan
smp purnama jakarta selatan
hendra prijatna
huruf dewanagari
smk bbc medan
sdit al hidayah
smp bina greha bogor
abc kids indramayu
budi asih
dr sasanti yuniar
sma negeri 1 porong
sdn neglasari sumedang
revenge r us
tk zubair bin awwam foto
pengertian himpunan kuasa
sds bhakti tugas
sedayu nusantara
smp insan mandiri bandar lampung
dokter tunjung wibowo
biaya sekolah an nahl jambi
sdit permata mojokerto
transport infoline
smk tambelang
dylan soedjasa
al wafa ciwidey
cahyo rahadian muzhar profil
logo sman 4 tambun selatan
smp 134
the right stuff
smp 8 padang
smpn 3 cisurupan
sman 1 teras
yayasan pendidikan insan pertiwi kabupaten bogor foto
sdit al-munawwar bogor foto
ras weddoid
sdit darul hijrah
smpit hayatan thayyibah
pkbm insan madani
tk alumna pekanbaru
sma bangun insan mandiri medan
smp negeri 2 kedungreja
silsilah kerajaan jerman
sdn sedati agung
sdn tanah tinggi 09
film sex barat
biaya masuk smp global insani tajur halang
sma negeri 1 juwana
stasiun hsr tegalluar
sd tahfidz ar rasyid cibinong
sma negeri 1 nalumsari
sdn tugu utara 13 pagi
nuril munfaridah
sman 1 muntok
smp trisula perwari 3
smkn 1 bangkinang
sman 47 jakarta
vera bahasuan profil
smkn 1 lelea
sma negeri 1 lohbener
kontemporer dewasa
smp 45 pekanbaru
sekolah daar en nisa bogor
smpn 2 mirit
Корень растения | Student Guru
Определение:
Корень — это подземный осевой вегетативный орган растения.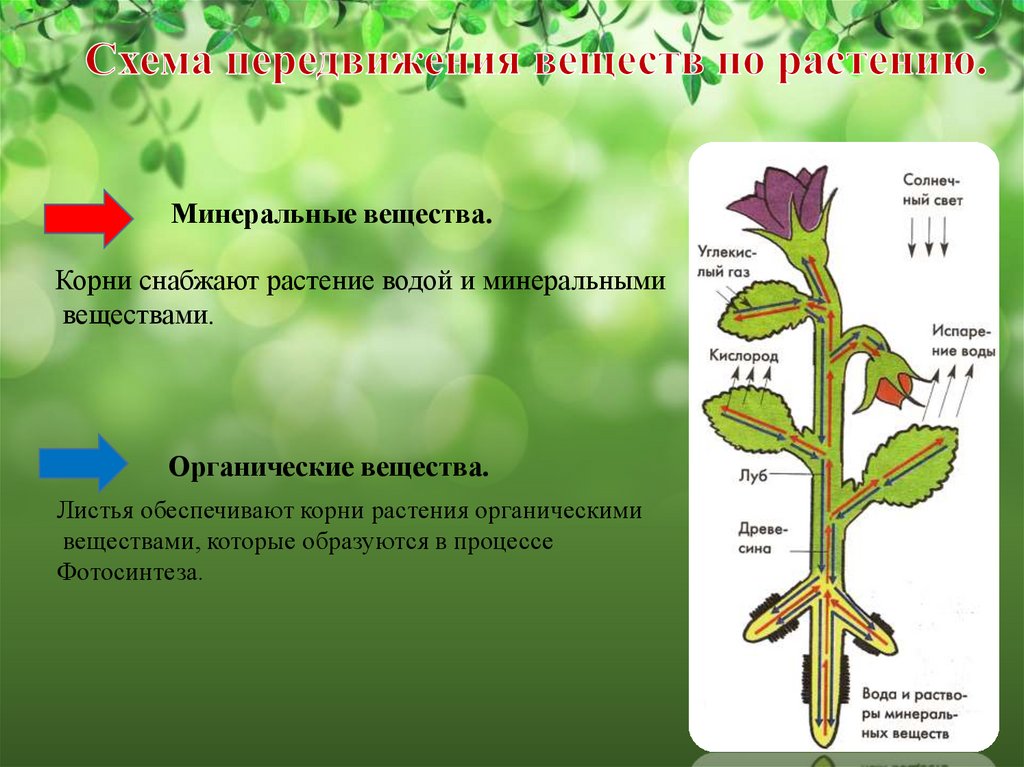 который имеет радиальную симметрию и обладает положительным геотропизмом. Корень, благодаря деятельности верхушечной меристемы, формирующей все его ткани, обладает способностью неограниченно долго нарастать в длину.
который имеет радиальную симметрию и обладает положительным геотропизмом. Корень, благодаря деятельности верхушечной меристемы, формирующей все его ткани, обладает способностью неограниченно долго нарастать в длину.
Функции корня растения.
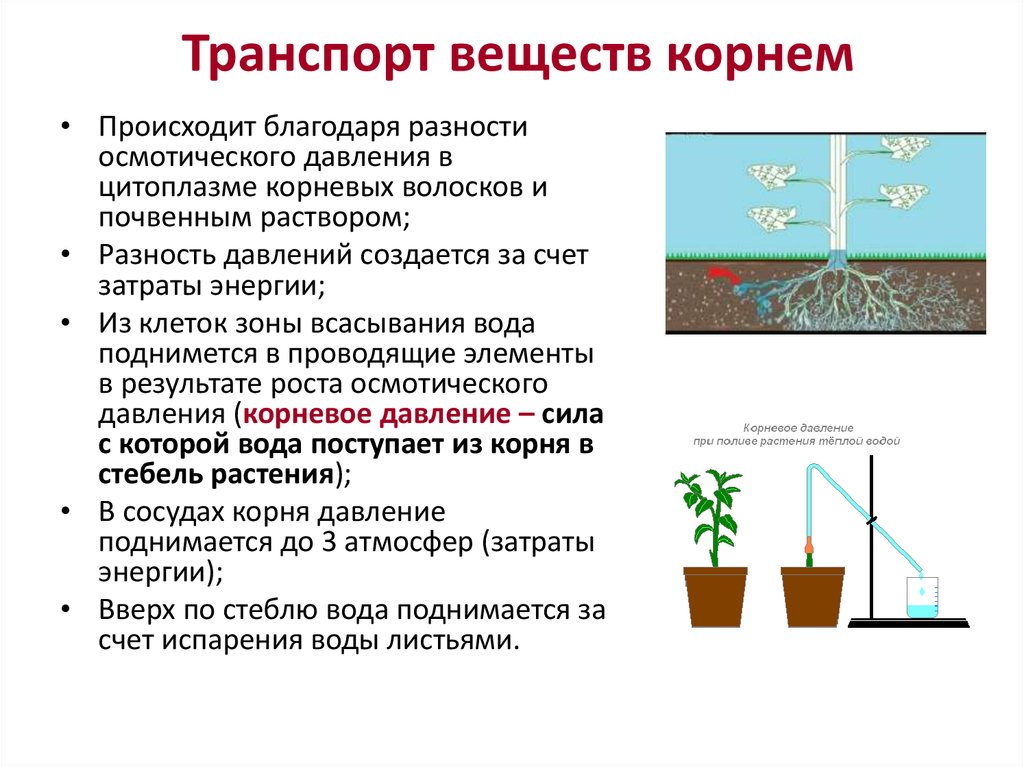
Основная функция корня растения — поглощение воды и минеральных веществ. Для выполнения этой функции необходимо, чтобы растение имело относительную неподвижность, т.е. было закреплено на каком-то определенном месте (опорная функция). Это становится возможным благодаря ветвлению корневой системы и ее положительному геотропизму. Геотропизм — ориентированный рост главного корня по направлению к центру Земли.
Благодаря корневым системам обеспечивается проведение растворов, так называемые, восходящий и нисходящий токи. Главным образом, это происходит под действием сил корневого давления и транспирации.
В результате первичного синтеза в корнях образуются аминокислоты, гормоны, ферменты и т. д. Все эти элементы быстро включаются в последующий биосинтез, который происходит в стебле и листьях растения. Таким образом, корень имеет метаболическое значение или еще можно сказать, что это функция синтеза биологически активных веществ.
д. Все эти элементы быстро включаются в последующий биосинтез, который происходит в стебле и листьях растения. Таким образом, корень имеет метаболическое значение или еще можно сказать, что это функция синтеза биологически активных веществ.
Также в корнях могут откладываться запасные питательные вещества, такие как крахмал, инулин и т.п. (Запасательная функция).
Для нормальной жизнедеятельности корневых систем необходимо
Благодаря адаптации к различным экологическим условиям у различных видов растений в процессе эволюции выработались особые функции — ходульные, сократительные и т.п. Корни могут взаимодействовать с грибами и микроорганизмами, живущими в почве, корнями других растений (микориза, клубеньки бобовых).
Морфология корня.
Корень является ведущим органом растения, и его формирование начинается в раннем онтогенезе организма. Поэтому уже на стадии прорастания семян с появления корневой системы начинается формирование проростка (см. рисунок ниже).
Поэтому уже на стадии прорастания семян с появления корневой системы начинается формирование проростка (см. рисунок ниже).
Корень формируется в раннем онтогенезе организма
Корневые системы растений.
Корневой системой называют совокупность всех корней одного растения. В нее входят: главный корень, боковые и придаточные корни. Типы корневых систем: корневые системы могут быть стержневыми или мочковатыми.
Виды корневых систем
При стержневой системе главный корень развивается из зародышевого корешка. Он составляет основу всей корневой системы и сохраняется пожизненно. Главный корень развивается в длину и толщину. Его легко отличить от других корней, т.к. он хорошо выделяется среди остальных корней. Кроме главного и боковых корней в стержневой корневой системе могут появляться и придаточные корни. Стержневая корневая система встречается у большинства двудольных и голосеменных растений.
Есть растения, у которых зародышевый корень живет недолго, следовательно их главный корень либо отмирает полностью, либо развит очень слабо. Их корневая система образуется из придаточных корней, которые вырастают у основания стебля, а на них возникают боковые корни. Такую корневую систему называют мочковатой. Такая корневая система встречается у всех однодольных растений и у некоторых двудольных, особенно у тех, которые размножаются вегетативно. У многих двудольных растений придаточные корни могут также образовываться на стеблях, засыпанных землей, или на подземных и ползучих стеблях.
Высшие споровые растения (к ним относятся папоротники, плауны, хвощи) не имеют главного корня. Он у них не образуется, т.к. заякоревание у этих растений слабое, а всасывание осуществляют или ризоиды, или придаточные корни, которые отходят от корневищ. Корневища этих растений — это видоизмененные стебли.
Развитие корневых систем сильно зависит от свойств почвы. Почва влияет на структуру корневой системы, на рост её корней, на глубину проникновения в почву и пространственное размещение корней в почве.
В почве вокруг корня растения выделяется ризосфера — зона богатая грибами, бактериями и другими микроорганизмами. Приспособление растений к условиям почвенного водоснабжения отражает то, как формируются поверхностные, глубинные и другие корневые системы.
В каждой корневой системе, кроме этого, происходят непрерывные изменения, которые связаны со сменой времен года, с возрастом растений и т.п.
Корень растет в длину неопределенно долго. Это происходит благодаря деятельности апикальной меристемы, ее нежные клетки практически всегда покрыты корневым чехликом. В отличие от побега у корня нет листьев, а, следовательно, отсутствуют расчленения на узлы и междоузлия. И в отличии от побега у корня присутствует чехлик. Длина растущей части корня не превышает 1 см.
Информация о статье:
Корень растения
Первая статья, описывающая корень растения. Включает в себя определение, функции и морфологию корня.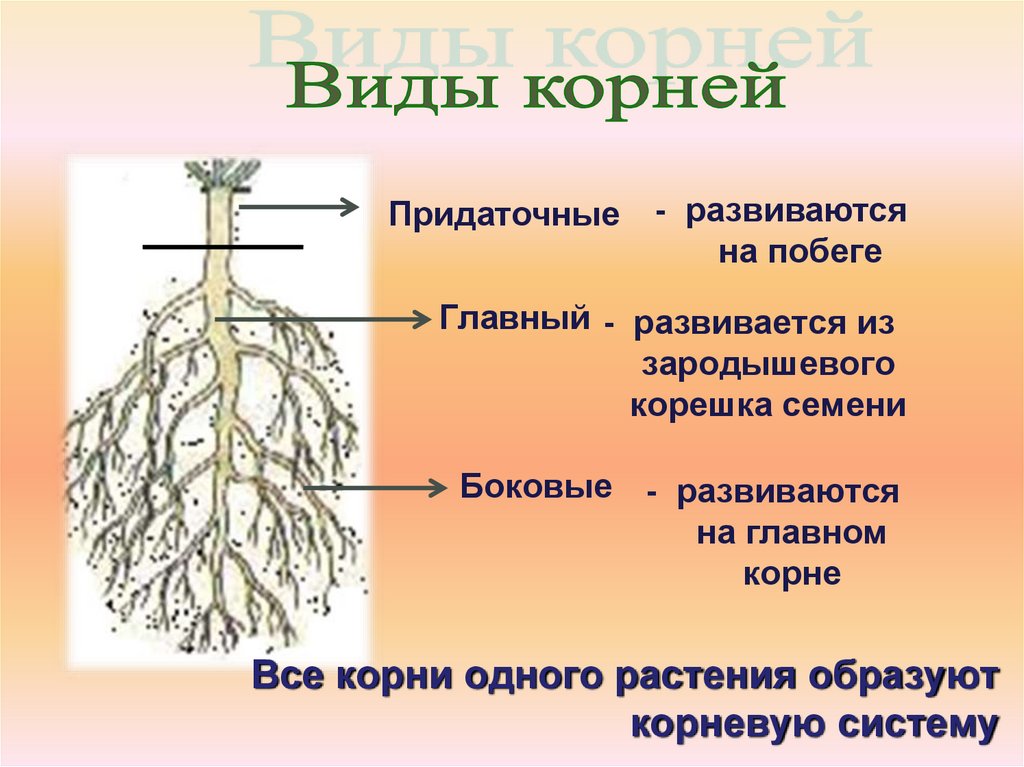
Written by: Stepan Gurov
Date Published: 12/09/2016
В статье описаны определения корня растений, функции корня, морфологическое строение корня. Описаны типы корневых систем растений.
10 / 10 stars
Перейти к оглавлению.
Закладка латерального корня у Arabidopsis thaliana: пробуждение силы Себастьян Дж., Боуман Дж.Л., Хелариутта Ю., Бенфей П.Н. Передача сигналов клетками с помощью микроРНК165/6 направляет зависящую от дозы гена судьбу клеток корня. Природа. 2010; 465:316–21. doi: 10.1038/nature08977. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/3133959
2. Hamant O, Heisler MG, Jonsson H, Krupinski P, Uyttewaal M, Bokov P, Corson F , Сахлин П., Будауд А., Мейеровиц Э.М., Кудер Ю., Траас Дж. Паттерн развития с помощью механических сигналов у арабидопсиса. Наука. 2008; 322:1650–5. doi: 10.1126/science.1165594. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/1145153
3. Heisenberg CP, Bellaiche Y. Силы в морфогенезе тканей и формировании паттерна. Клетка. 2013; 153:948–62. doi: 10.1016/j.cell.2013.05.008. [PubMed] [CrossRef] [Google Scholar]
Heisenberg CP, Bellaiche Y. Силы в морфогенезе тканей и формировании паттерна. Клетка. 2013; 153:948–62. doi: 10.1016/j.cell.2013.05.008. [PubMed] [CrossRef] [Google Scholar]
4. Lavenus J, Goh T, Roberts I, Guyomarc’h S, Lucas M, De Smet I, Fukaki H, Beeckman T, Bennett M, Laplaze L. Развитие латерального корня у арабидопсиса: пятьдесят оттенков ауксина. Тенденции Растениевод. 2013;18:450–8. doi: 10.1016/j.tplants.2013.04.006. [PubMed] [CrossRef] [Академия Google]
5. Schlereth A, Moller B, Liu W, Kientz M, Flipse J, Rademacher EH, Schmid M, Jurgens G, Weijers D. MONOPTEROS контролирует инициацию эмбрионального корня, регулируя подвижный транскрипционный фактор. Природа. 2010; 464:913–6. doi: 10.1038/nature08836. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/2478966
6. Monshausen GB, Haswell ES. Сила природы: молекулярные механизмы механовосприятия у растений. J Опытный бот. 2013;64:4663–80. doi: 10.1093/jxb/ert204. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
7. Nakayama N, Smith RS, Mandel T, Robinson S, Kimura S, Boudaoud A, Kuhlemeier C. Механическая регуляция роста, опосредованного ауксином. Карр Биол. 2012; 22:1468–76. doi: 10.1016/j.cub.2012.06.050. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/717952919
Nakayama N, Smith RS, Mandel T, Robinson S, Kimura S, Boudaoud A, Kuhlemeier C. Механическая регуляция роста, опосредованного ауксином. Карр Биол. 2012; 22:1468–76. doi: 10.1016/j.cub.2012.06.050. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/717952919
8. Boudaoud A. Введение в механику морфогенеза для биологов растений. Тенденции Растениевод. 2010;15:353–60. doi: 10.1016/j.tplants.2010.04.002. [PubMed] [CrossRef] [Google Scholar]
9. Sampathkumar A, Krupinski P, Wightman R, Milani P, Berquand A, Boudaoud A, Hamant O, Jonsson H, Meyerowitz EM. Субклеточный и надклеточный механический стресс определяет поведение цитоскелета в клетках мостовой семядолей арабидопсиса. Элиф. 2014;3:e01967. doi: 10.7554/eLife.01967. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
10. Уюттевал М., Буриан А., Алим К., Ландрейн Б., Боровска-Викрет Д., Дедье А., Посель А., Лудиния М., Траас Дж., Будауд А. , Квятковска Д., Хамант О. Механический стресс действует через катанин, усиливая различия в скорости роста между соседними клетками арабидопсиса. Клетка. 2012; 149: 439–51. doi: 10.1016/j.cell.2012.02.048. [PubMed] [CrossRef] [Google Scholar]
Клетка. 2012; 149: 439–51. doi: 10.1016/j.cell.2012.02.048. [PubMed] [CrossRef] [Google Scholar]
11. Лукас М., Кеноби К., фон Вангенхайм Д., Вобета У., Сваруп К., Де Смет И., Ван Дамм Д., Лоуренс Т., Перет Б., Москарди Э., Барбо Д., Годин С., Солт Д., Гайомарх С., Стельцер Э. Х., Майзель А., Лаплаз Л., Беннетт М.Дж. Морфогенез боковых корней зависит от механических свойств покрывающих тканей. Proc Natl Acad Sci USA. 2013;110:5229–34. doi: 10.1073/pnas.1210807110. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/718019793
12. Richter GL, Monshausen GB, Krol A, Gilroy S. Механические стимулы модулируют органогенез боковых корней. Завод Физиол. 2009; 151:1855–66. doi: 10.1104/стр.109.142448. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
13. Сваруп К., Бенкова Э., Сваруп Р., Касимиро И., Перет Б., Ян И., Парри Г., Нильсен Э., Де Смет И., Ваннесте С., Levesque MP, Carrier D, James N, Calvo V, Ljung K, Kramer E, Roberts R, Graham N, Marillonnet S, Patel K, Jones JD, Taylor CG, Schachtman DP, May S, Sandberg G, Benfey P, Friml J , Керр И.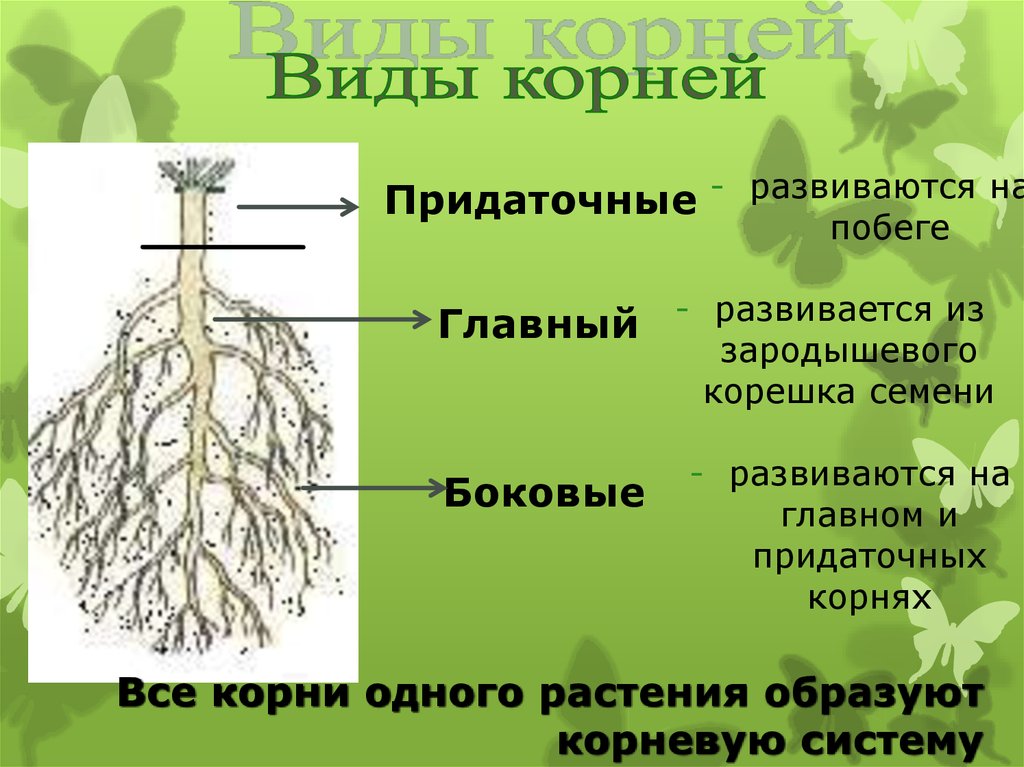 , Бекман Т., Лаплаз Л., Беннетт М.Дж. Переносчик притока ауксина LAX3 способствует появлению боковых корней. Nat Cell Biol. 2008;10:946–54. дои: 10.1038/ncb1754. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/1120929
, Бекман Т., Лаплаз Л., Беннетт М.Дж. Переносчик притока ауксина LAX3 способствует появлению боковых корней. Nat Cell Biol. 2008;10:946–54. дои: 10.1038/ncb1754. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/1120929
14. Vermeer JE, von Wangenheim D, Barberon M, Lee Y, Stelzer EH, Maizel A, Geldner N. Пространственное приспособление соседними клетками требуется для инициации органов у Arabidopsis. Наука. 2014; 343:178–83. doi: 10.1126/science.1245871. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/718233332
15. Peret B, De Rybel B, Casimiro I, Benkova E, Swarup R, Laplaze L, Beeckman T, Bennett MJ. Развитие боковых корней арабидопсиса: новая история. Тенденции Растениевод. 2009 г.;14:399–408. doi: 10.1016/j.tplants.2009.05.002. [PubMed] [CrossRef] [Google Scholar]
16. Вилчес-Барро А., Майзель А. Разговор сквозь стены: механизмы появления боковых корней у Arabidopsis thaliana. Curr Opin Plant Biol. 2014; 23:31–38. doi: 10.1016/j.pbi.2014.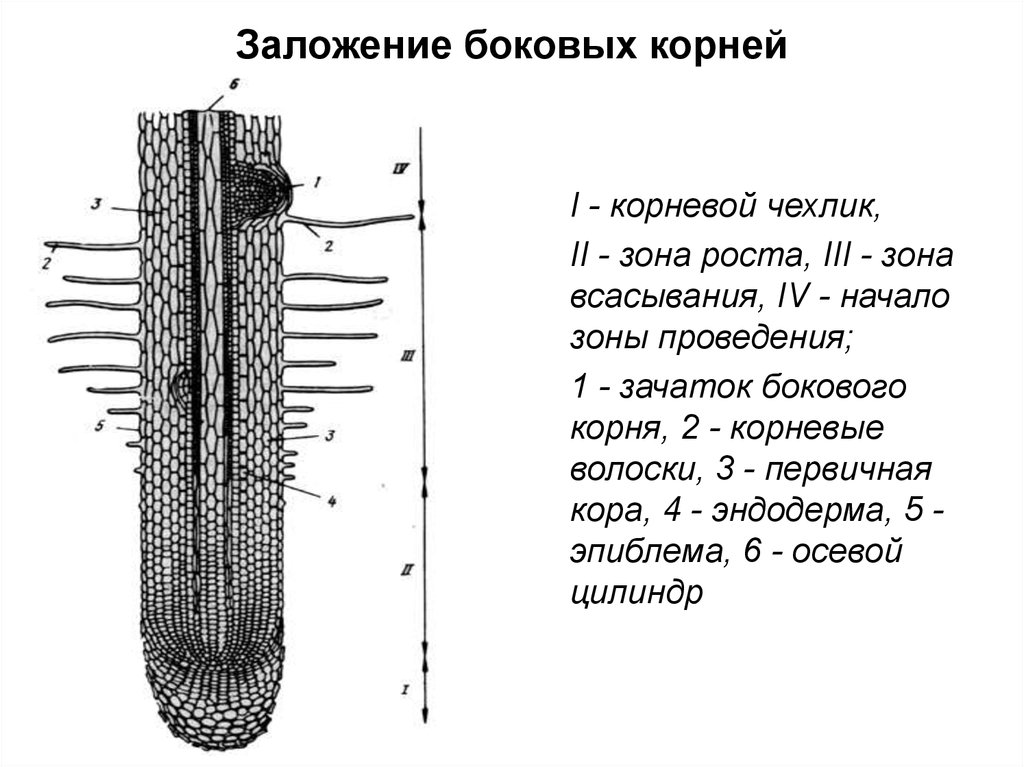 10.005. [PubMed] [CrossRef] [Google Scholar]
10.005. [PubMed] [CrossRef] [Google Scholar]
17. Kumpf RP, Shi CL, Larrieu A, Sto IM, Butenko MA, Peret B, Riiser ES, Bennett MJ, Aalen RB. Пептид абсциссии цветочных органов IDA и его рецепторы HAE/HSL2 контролируют разделение клеток во время появления боковых корней. Proc Natl Acad Sci USA. 2013;110:5235–40. doi: 10.1073/pnas.1210835110. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
18. Перет Б., Ли Г., Чжао Дж., Бэнд Л.Р., Восс Ю., Постер О., Луу Д.Т., Да Инес О., Касимиро И., Лукас М., Уэллс Д.М., Лаззерини Л., Накри П., Кинг Дж.Р., Дженсен О.Е., Шаффнер А.Р., Морел С., Беннетт М.Дж. Ауксин регулирует функцию аквапоринов, облегчая появление боковых корней. Натт Селл Биол. 2012;14:991–8. дои: 10.1038/ncb2573. [PubMed] [CrossRef] [Google Scholar]
19. Parizot B, Laplaze L, Ricaud L, Boucheron-Dubuisson E, Bayle V, Bonke M, De Smet I, Poethig SR, Helariutta Y, Haseloff J, Chriqui D, Beeckman T, Nussaume L. Диархическая симметрия сосудистого пучка в корне Arabidopsis охватывает перицикл и отражается в закладке двустихного латерального корня.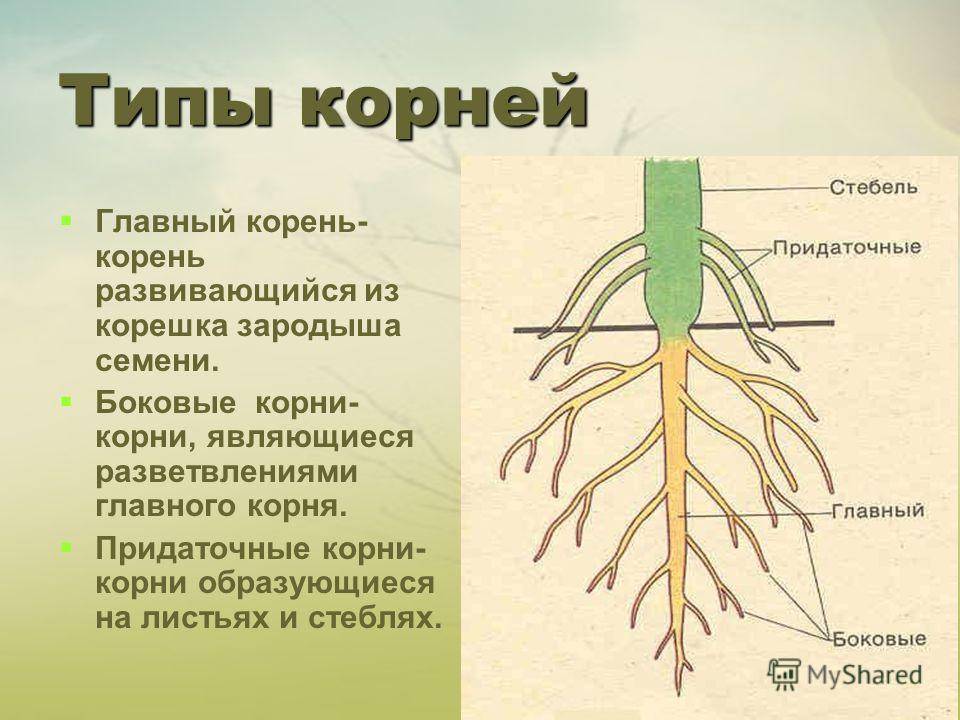 Завод Физиол. 2008; 146:140–8. doi: 10.1104/стр.107.107870. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/1099314
Завод Физиол. 2008; 146:140–8. doi: 10.1104/стр.107.107870. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/1099314
20. Ван Норман Дж.М., Суан В., Бекман Т., Бенфи П.Н. Ветвиться или не ветвиться: роль предварительного рисунка в формировании боковых корней. Разработка. 2013; 140:4301–10. doi: 10.1242/dev.090548. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
21. Де Смет И., Тецумура Т., Де Рибель Б., Фрей Н.Ф., Лаплаз Л., Касимиро И., Сваруп Р., Наудтс М., Ваннесте С., Оденарт Д. , Inze D, Bennett MJ, Beeckman T. Ауксин-зависимая регуляция положения боковых корней в базальной меристеме арабидопсиса. Разработка. 2007; 134: 681–9.0. doi: 10.1242/dev.02753. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/1060733
22. Морено-Рисуэно М.А., Ван Норман Дж.М., Морено А., Чжан Дж., Анерт С.Е., Бенфей П.Н. Осциллирующая экспрессия генов определяет способность к периодическому ветвлению корней арабидопсиса. Наука.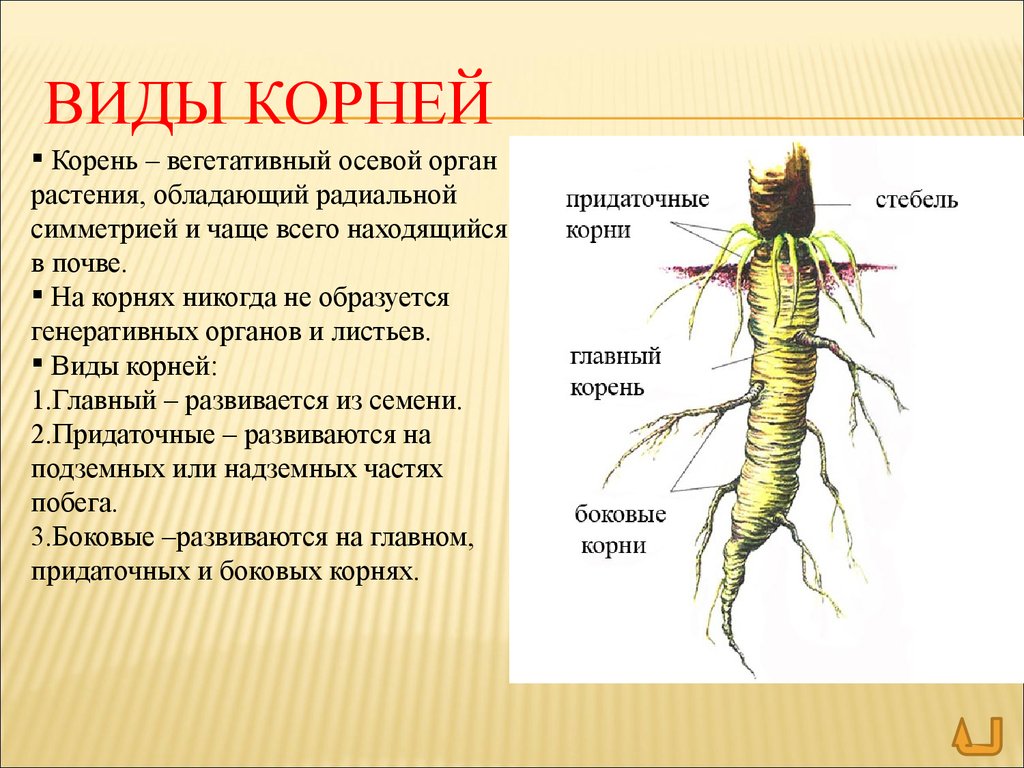 2010;329:1306–11. doi: 10.1126/science.1191937. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/5235956
2010;329:1306–11. doi: 10.1126/science.1191937. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/5235956
23. Van Norman JM, Zhang J, Cazzonelli CI, Pogson BJ, Harrison PJ, Bugg TD, Chan KX, Томпсон А.Дж., Бенфи П.Н. Периодическое ветвление корня у арабидопсиса требует синтеза неохарактеризованного каротиноидного производного. Proc Natl Acad Sci USA. 2014;111:E1300. doi: 10.1073/pnas.1403016111. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/718313253
24. Улмасов Т., Мерфетт Дж., Хаген Г., Гилфойл Т.Дж. Белки Aux/IAA подавляют экспрессию репортерных генов, содержащих природные и высокоактивные синтетические элементы ответа на ауксин. Растительная клетка. 1997; 9: 1963–71. doi: 10.1105/tpc.9.11.1963. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
25. Lokerse AS, Weijers D. Auxin входит в матрицу-сборку механизмов реагирования для конкретных результатов. Текущее мнение Раст. биол.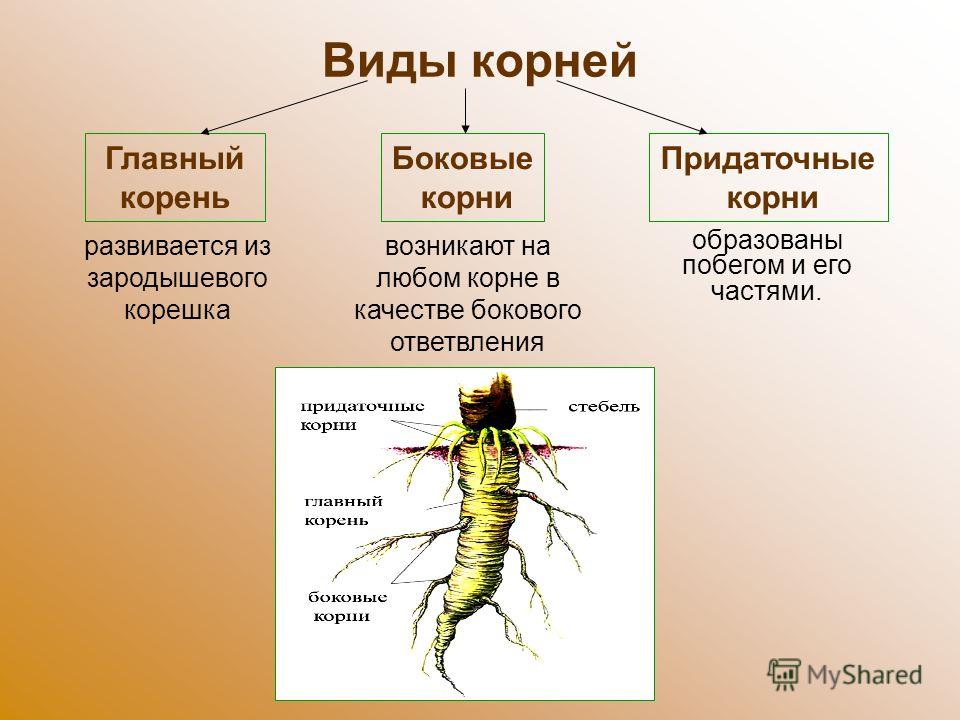 2009;12:520–6. doi: 10.1016/j.pbi.2009.07.007. [PubMed] [CrossRef] [Академия Google]
2009;12:520–6. doi: 10.1016/j.pbi.2009.07.007. [PubMed] [CrossRef] [Академия Google]
26. Де Рибель Б., Васильева В., Паризо Б., Демелэнэр М., Грюневальд В., Ауденарт Д., Ван Кампенхаут Дж., Овервурде П., Янсен Л., Ваннесте С., Моллер Б., Уилсон М., Холман Т., Ван Истердал Г., Брюно G, Vuylsteke M, Vernoux T, De Veylder L, Inze D, Weijers D, Bennett MJ, Beeckman T. Новый сигнальный каскад aux/IAA28 активирует GATA23-зависимую спецификацию идентичности клеток-основателей боковых корней. Карр Биол. 2010;20:1697–706. doi: 10.1016/j.cub.2010.09.007. [PubMed] [CrossRef] [Академия Google]
27. Де Смет И., Лау С., Восс У., Ваннесте С., Бенджаминс Р., Радемахер Э.Х., Шлерет А., Де Рибель Б., Васильева В., Грюневальд В., Наудтс М., Левеск М.П., Эхрисманн Дж.С., Инзе Д., Люшниг С. , Benfey PN, Weijers D, Van Montagu MC, Bennett MJ, Jurgens G, Beeckman T. Бимодулярный ответ ауксина контролирует органогенез у арабидопсиса. Proc Natl Acad Sci USA. 2010;107:2705–10. doi: 10.1073/pnas.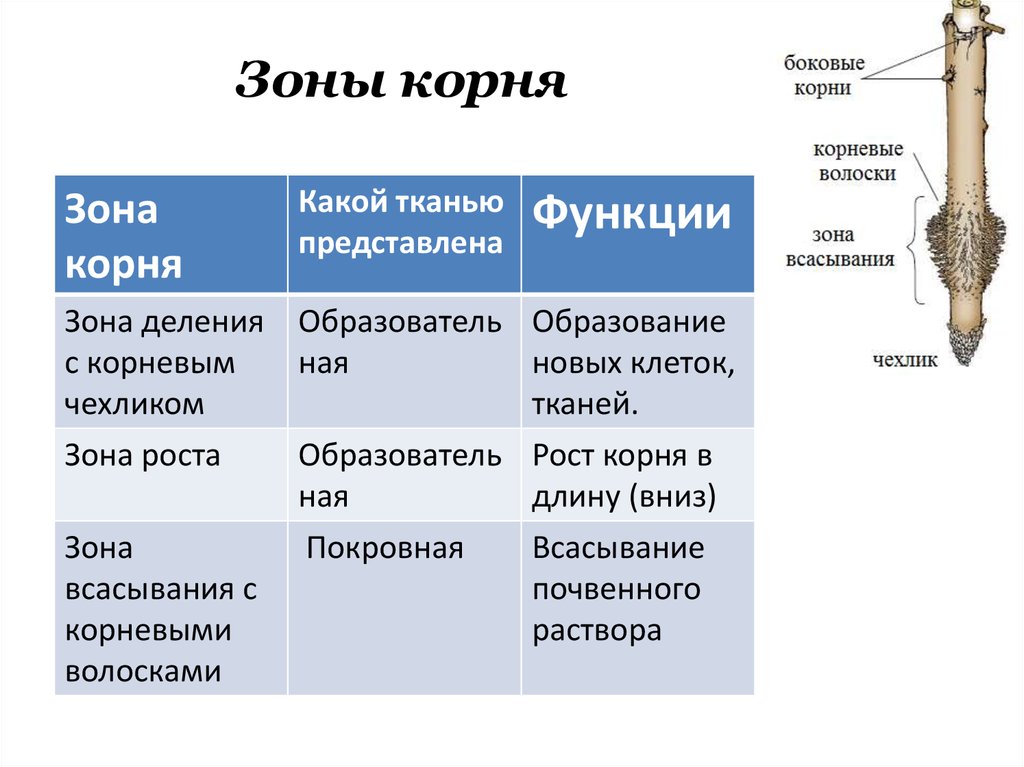 0915001107. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
0915001107. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
28. Goh T, Kasahara H, Mimura T, Kamiya Y, Fukaki H. Несколько модулей AUX/IAA-ARF регулируют формирование боковых корней: роль арабидопсиса SHY2/IAA3-опосредованная передача сигналов ауксина. Philos Trans R Soc Lond B Biol Sci. 2012; 367:1461–8. дои: 10.1098/рстб.2011.0232. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
29. Okushima Y, Fukaki H, Onoda M, Theologis A, Tasaka M. ARF7 и ARF19 регулируют формирование боковых корней путем прямой активации генов LBD/ASL у Арабидопсис. Растительная клетка. 2007; 19: 118–30. doi: 10.1105/tpc.106.047761. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
30. Bielach A, Podlesakova K, Marhavy P, Duclercq J, Cuesta C, Muller B, Grunewald W, Tarkowski P, Benkova E. Пространственно-временная регуляция латеральной Органогенез корня арабидопсиса с помощью цитокинина. Растительная клетка. 2012;24:3967–81. doi: 10.1105/tpc.112.103044. [Статья бесплатно PMC] [PubMed] [CrossRef] [Google Scholar]
31. Чанг Л., Рамиредди Э., Шмуллинг Т. Образование боковых корней и рост арабидопсиса избыточно регулируются метаболизмом цитокининов и сигнальными генами. J Опытный бот. 2013;64:5021–32. doi: 10.1093/jxb/ert291. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Чанг Л., Рамиредди Э., Шмуллинг Т. Образование боковых корней и рост арабидопсиса избыточно регулируются метаболизмом цитокининов и сигнальными генами. J Опытный бот. 2013;64:5021–32. doi: 10.1093/jxb/ert291. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
32. Laplaze L, Benkova E, Casimiro I, Maes L, Vanneste S, Swarup R, Weijers D, Calvo V, Parizot B, Herrera-Rodriguez MB , Offringa R, Graham N, Doumas P, Friml J, Bogusz D, Beeckman T, Bennett M. Цитокинины действуют непосредственно на клетки-основатели боковых корней, подавляя закладку корней. Растительная клетка. 2007;19: 3889–900. doi: 10.1105/tpc.107.055863. [PMC бесплатная статья] [PubMed] [CrossRef] [Google Scholar]
33. Marhavy P, Duclercq J, Weller B, Feraru E, Bielach A, Offringa R, Friml J, Schwechheimer C, Murphy A, Benkova E. Цитокинин контролирует полярность PIN1-зависимого транспорта ауксина во время органогенеза латерального корня. Карр Биол. 2014; 24:1031–1037. doi: 10.1016/j.cub.2014.04.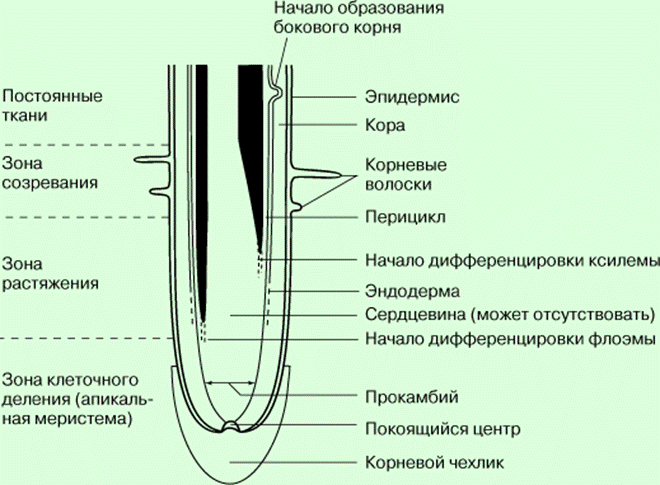 002. [PubMed] [CrossRef] [Google Scholar]
002. [PubMed] [CrossRef] [Google Scholar]
34. Goh T, Joi S, Mimura T, Fukaki H. Установление асимметрии в клетках-основателях боковых корней арабидопсиса регулируется LBD16/ASL18 и родственными LBD/ASL белками. Разработка. 2012;139: 883–93. doi: 10.1242/dev.071928. [PubMed] [CrossRef] [Google Scholar]
35. Tamura K, Iwabuchi K, Fukao Y, Kondo M, Okamoto K, Ueda H, Nishimura M, Hara-Nishimura I. Миозин XI-i связывает ядерную мембрану с цитоскелет, контролирующий движение и форму ядер у арабидопсиса. Карр Биол. 2013; 23:1776–81. doi: 10.1016/j.cub.2013.07.035. [PubMed] [CrossRef] [Google Scholar]
36. Marhavy P, Vanstraelen M, De Rybel B, Zhaojun D, Bennett MJ, Beeckman T, Benkova E. Рефлюкс ауксина между энтодермой и перициклом способствует инициации латерального корня. EMBO Дж. 2013; 32:149–58. doi: 10.1038/emboj.2012.303. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. Lee Y, Rubio MC, Alassimone J, Geldner N. Механизм локализованного отложения лигнина в эндодерме.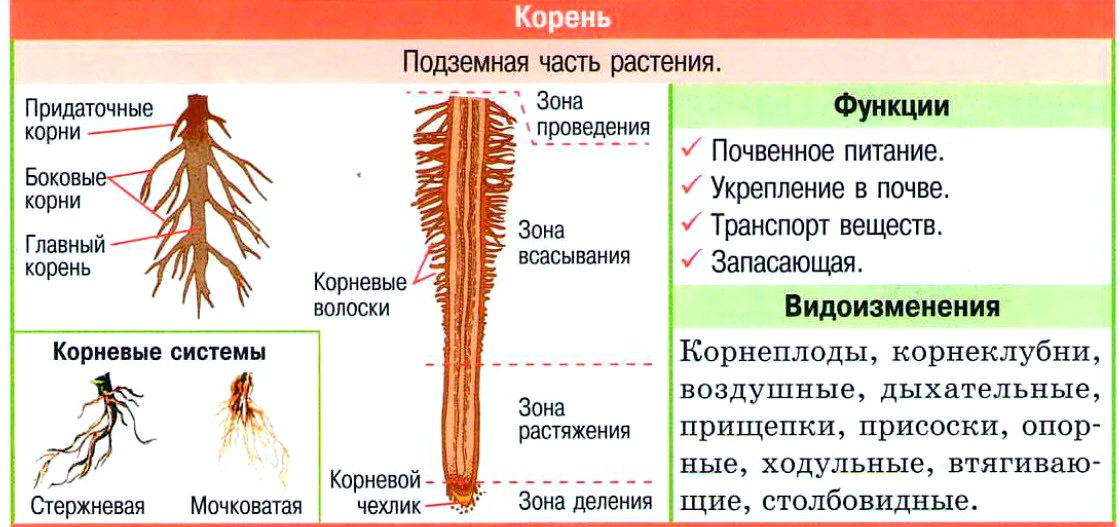 Клетка. 2013; 153:402–12. doi: 10.1016/j.cell.2013.02.045. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/718002366
Клетка. 2013; 153:402–12. doi: 10.1016/j.cell.2013.02.045. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/718002366
38. Naseer S, Lee Y, Lapierre C, Franke R, Nawrath C, Geldner N. Диффузионный барьер полосы Каспариана у арабидопсиса из полимера лигнина без суберина. Proc Natl Acad Sci USA. 2012;109: 10101–6. doi: 10.1073/pnas.1205726109. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
39. Roppolo D, De Rybel B, Tendon VD, Pfister A, Alassimone J, Vermeer JE, Yamazaki M, Stierhof YD, Beeckman T, Geldner N. Новое семейство белков обеспечивает образование полосы Каспари в энтодерме. Природа. 2011; 473:380–3. doi: 10.1038/nature10070. [PubMed] [CrossRef] [Google Scholar]
40. Bao Y, Aggarwal P, Robbins NE, 2nd, Sturrock CJ, Thompson MC, Tan HQ, Tham C, Duan L, Rodriguez PL, Vernoux T, Mooney SJ, Bennett MJ, Диннени JR. Корни растений используют механизм формирования паттерна для размещения боковых корневых ветвей по направлению к доступной воде.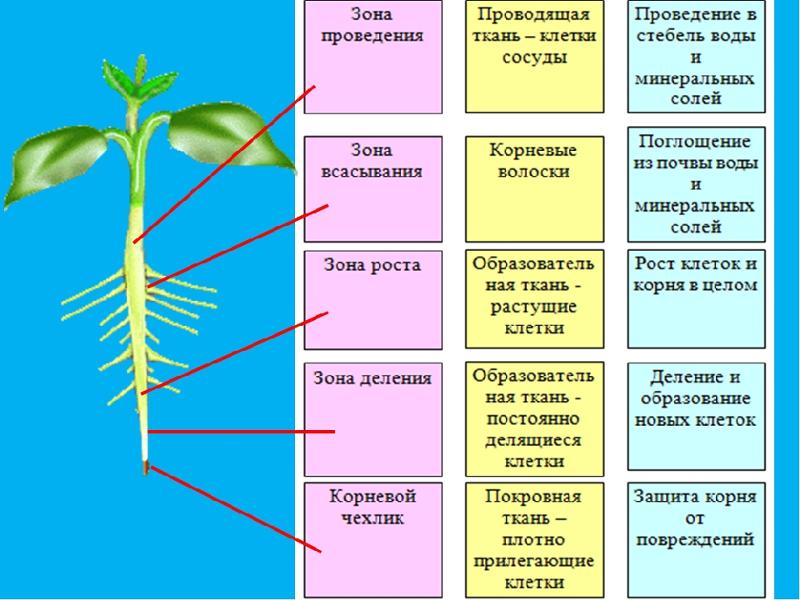 Proc Natl Acad Sci USA. 2014;111:9319–24. doi: 10.1073/pnas.1400966111. [PMC бесплатная статья] [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/718448127
Proc Natl Acad Sci USA. 2014;111:9319–24. doi: 10.1073/pnas.1400966111. [PMC бесплатная статья] [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/718448127
41. Beauzamy L, Nakayama N, Boudaoud A. Цветы под давлением: тонкости регулирования тургора в разработка. Энн Бот. 2014; 114:1517–33. doi: 10.1093/aob/mcu187. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
42. Vaten A, Dettmer J, Wu S, Stierhof YD, Miyashima S, Yadav SR, Roberts CJ, Campilho A, Bulone V, Lichtenberger R, Lehesranta S, Mahonen AP, Kim JY, Jokitalo E, Sauer N, Scheres B, Nakajima K, Carlsbecker A, Gallagher KL, Helariutta Y. Биосинтез каллозы регулирует симпластический транспорт во время развития корня. Ячейка Дев. 2011;21:1144–55. doi: 10.1016/j.devcel.2011.10.006. [PubMed] [CrossRef] [Академия Google] http://f1000.com/prime/13408956
43. Бенитес-Альфонсо Й., Фолкнер С., Пендл А., Мияшима С., Хелариутта Й., Мауле А. Симпластическая межклеточная связь регулирует формирование боковых корней.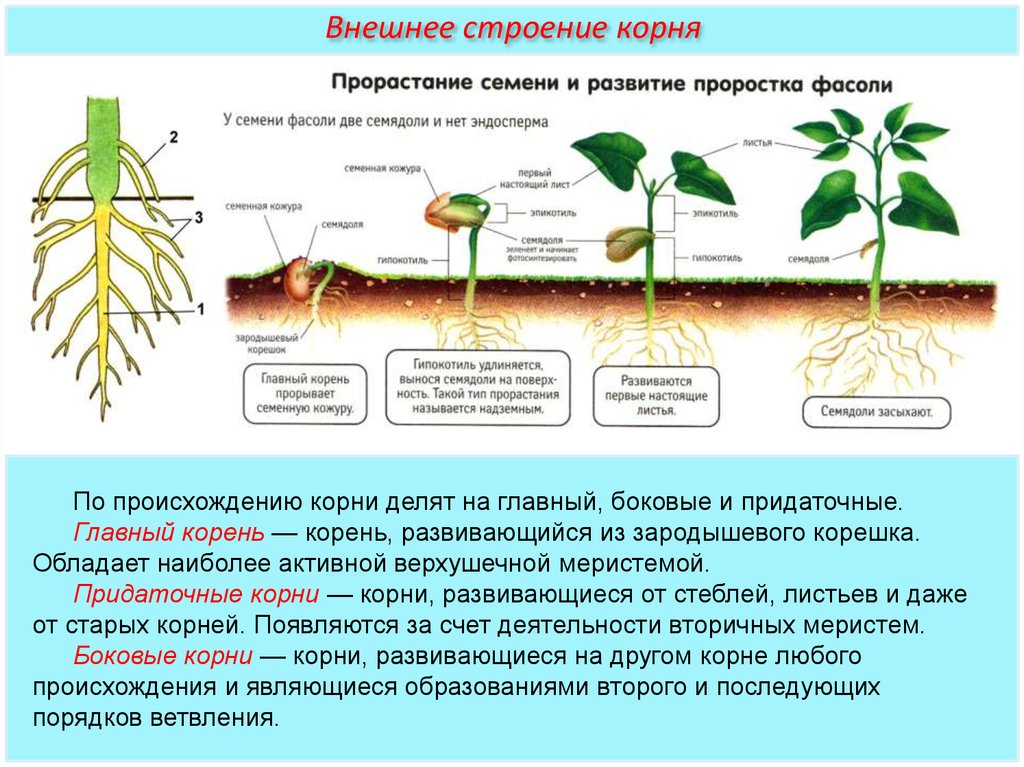 Ячейка Дев. 2013;26:136–47. doi: 10.1016/j.devcel.2013.06.010. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/718030982
Ячейка Дев. 2013;26:136–47. doi: 10.1016/j.devcel.2013.06.010. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/718030982
44. Ellinger D, Voigt CA. Биосинтез каллозы у арабидопсиса с акцентом на реакцию патогенов: что мы узнали за последнее десятилетие. Энн Бот. 2014; 114:1349–58. doi: 10.1093/aob/mcu120. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
45. Ditengou FA, Teale WD, Kochersperger P, Flittner KA, Kneuper I, van der Graaff E, Nziengui H, Pinosa F, Li X, Nitschke R, Laux T, Palme K. Механическая индукция закладки боковых корней у арабидопсиса Талиана. Proc Natl Acad Sci USA. 2008; 105:18818–23. doi: 10.1073/pnas.0807814105. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
46. Vincill ED, Clarin AE, Molenda JN, Spalding EP. Взаимодействующие белки, подобные глутаматным рецепторам, во флоэме регулируют инициацию боковых корней у арабидопсиса. Растительная клетка. 2013; 25:1304–13. doi: 10.1105/tpc.113.110668. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] http://f1000. com/prime/718001374
com/prime/718001374
47. Haswell ES, Peyronnet R, Barbier-Brygoo H, Meyerowitz EM, Frachisse JM. Два гомолога MscS обеспечивают активность механочувствительных каналов в корне арабидопсиса. Карр Биол. 2008;18:730–4. doi: 10.1016/j.cub.2008.04.039. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/1123013
48. Yuan F, Yang H, Xue Y, Kong D, Ye R, Li C, Zhang J, Theprungsirikul L, Shrift Т., Кричильский Б., Джонсон Д.М., Свифт Г.Б., Хе Ю., Сидоу Д.Н., Пей З.М. OSCA1 опосредует вызванное осмотическим стрессом увеличение содержания Ca2+, жизненно важное значение для осмочувствительности у арабидопсиса. Природа. 2014; 514:367–71. дои: 10.1038/nature13593. [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/718546337
49. Basssel GW, Stamm P, Mosca G, Barbier de Reuille P, Gibbs DJ, Winter R, Janka A, Холдсворт М.Дж., Смит Р.С. Механические ограничения, налагаемые трехмерной клеточной геометрией и расположением, модулируют модели роста эмбриона арабидопсиса. Proc Natl Acad Sci USA. 2014; 111:8685–90. doi: 10.1073/pnas.1404616111. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Proc Natl Acad Sci USA. 2014; 111:8685–90. doi: 10.1073/pnas.1404616111. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
50. Yoshida S, Barbier de Reuille P, Lane B, Basssel GW, Prusinkiewicz P, Smith RS, Weijers D. Генетический контроль развития растений путем переопределения Правило геометрического деления. Ячейка Дев. 2014;29: 75–87. doi: 10.1016/j.devcel.2014.02.002. [PubMed] [CrossRef] [Google Scholar]
51. Marin E, Jouannet V, Herz A, Lokerse AS, Weijers D, Vaucheret H, Nussaume L, Crespi MD, Maizel A. miR390, тасиРНК TAS3 Arabidopsis и их ауксин Цели RESPONSE FACTOR определяют ауторегуляторную сеть, количественно регулирующую рост боковых корней. Растительная клетка. 2010;22:1104–17. doi: 10.1105/tpc.109.072553. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] http://f1000.com/prime/2910962
Развитие растений II: первичный и вторичный рост
Цели обучения
- Различать первичный и вторичный рост
- Определите и опишите роль верхушечных и боковых меристем в росте растений
- Сравните и сопоставьте процессы и результаты первичного и вторичного роста стеблей и корней
- Опишите функцию и организацию одревесневших стеблей, полученных в результате вторичного роста
Индетерминантный рост растений: меристемы
Приведенная ниже информация была адаптирована из OpenStax Biology 30. 1
1
В отличие от большинства животных, которые вырастают до определенного размера и формы тела, а затем перестают расти ( детерминированный рост), растения демонстрируют неопределенный рост, при котором растение будет продолжать добавлять новые органы (листья, стебли, корни) до тех пор, пока у него есть доступ к необходимым ресурсам. Растения могут продолжать расти бесконечно благодаря специализированным тканям, называемым меристемами , которые являются областями непрерывного деления и роста клеток. Клетки меристематической ткани либо недифференцированы, либо не полностью дифференцированы, и они продолжают продуцировать клетки, которые быстро дифференцируются, или специализируются и становятся постоянными тканями (дермальными, основными и сосудистыми).
Меристематические ткани делятся на три типа в зависимости от их расположения в растении. Верхушечные меристемы содержат меристематическую ткань, расположенную на концах стеблей и корней, которая позволяет растению вытягиваться в длину.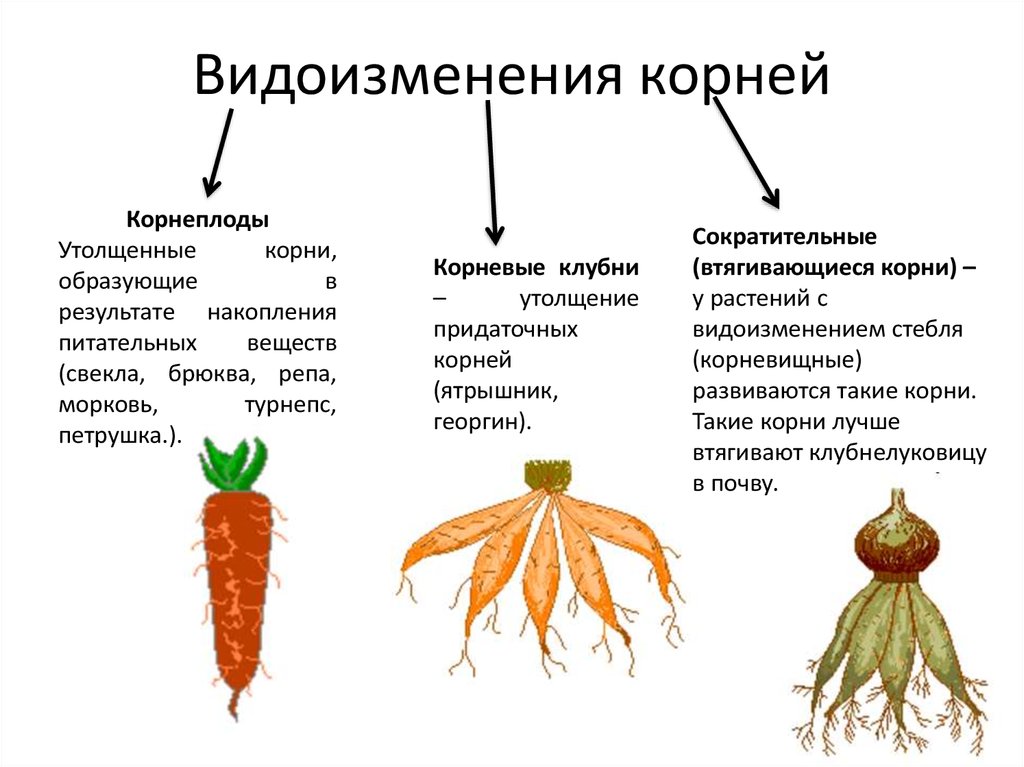 Боковые меристемы облегчают рост в толщине или обхвате у созревающего растения. Интеркалярные (также называемые базальными ) меристемы встречаются только у некоторых однодольных, в основании листовых пластинок и в узлах (областях, где листья прикрепляются к стеблю). Эта ткань позволяет листовой пластинке однодольного растения увеличиваться в длину от основания листа; например, он позволяет листьям газонной травы удлиняться даже после многократного выпаса скота или скашивания.
Боковые меристемы облегчают рост в толщине или обхвате у созревающего растения. Интеркалярные (также называемые базальными ) меристемы встречаются только у некоторых однодольных, в основании листовых пластинок и в узлах (областях, где листья прикрепляются к стеблю). Эта ткань позволяет листовой пластинке однодольного растения увеличиваться в длину от основания листа; например, он позволяет листьям газонной травы удлиняться даже после многократного выпаса скота или скашивания.
Меристемы способствуют как первичному (выше/длиннее), так и вторичному (шире) росту. Первичный рост контролируется апикальными меристемами корня или апикальными меристемами побега , а вторичный рост контролируется двумя боковыми меристемами, называемыми сосудистым камбием и пробковым камбием . Не все растения проявляют вторичный рост.
В приведенном ниже видеоролике подробно рассказывается о первичном и вторичном росте растений (начало с 2:20):
Первичный рост корней
Приведенная ниже информация была адаптирована из OpenStax Biology 30.3
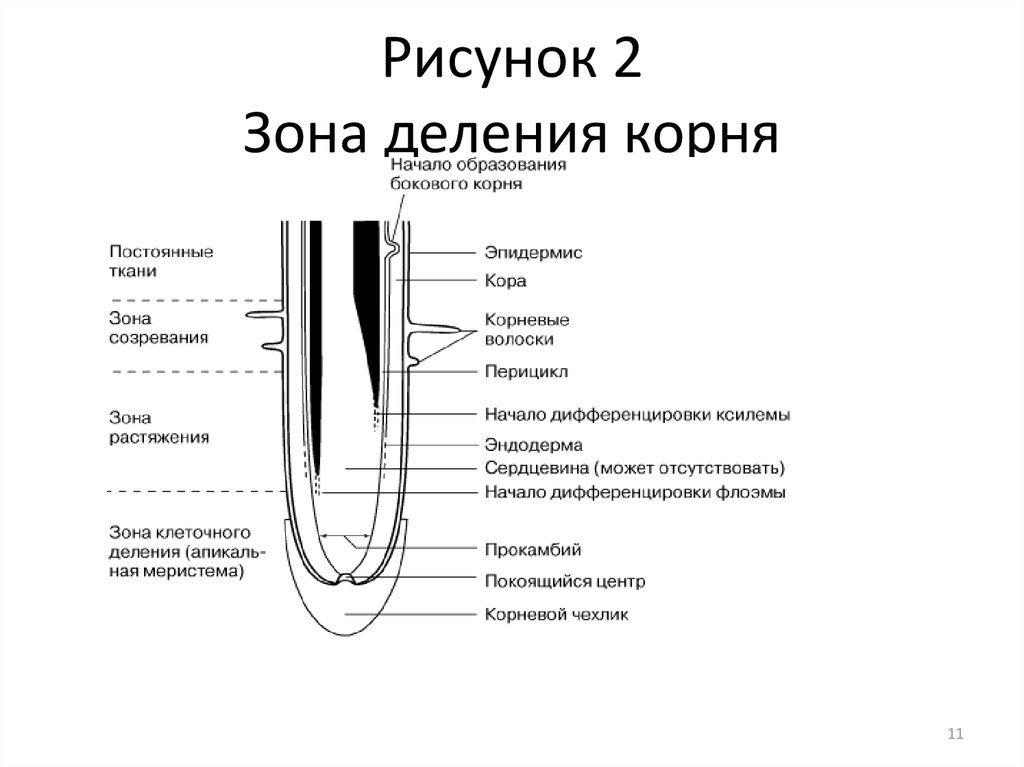
Рост корней начинается с прорастания семян. Когда зародыш растения выходит из семени, корешок зародыша образует корневую систему. Кончик корня защищен корневым чехликом , структурой, эксклюзивной для корней и не похожей ни на одну другую структуру растений. Корневой чехлик постоянно заменяется, потому что он легко повреждается, когда корень проталкивается через почву. За корневым чехликом, в пределах первого сантиметра или около того, кончик корня можно разделить на три зоны:
- Зона клеточного деления , которая содержит апикальную меристему, представляет собой место непосредственно за корневым чехликом, где клетки активно делятся путем митоза.

- Зона клеточного удлинения представляет собой место, где новообразованные клетки растут или увеличиваются в длину, увеличивая длину корня. Этот процесс требует поглощения воды, которая буквально растягивает клетки и увеличивает их размер.
- Зона клеточного созревания – это место, где вновь удлиненные клетки завершают свою дифференцировку в дермальную, сосудистую или основную ткани. Созревание обусловлено изменениями в экспрессии генов.

Кончик корня делится на три зоны: верхнюю зону созревания, среднюю зону удлинения и нижнюю зону деления клеток на кончике корня. В области созревания от основного корня отходят корневые волоски, клетки крупные и прямоугольные. Участок удлинения не имеет корневых волосков, а клетки все еще прямоугольные, но несколько мельче. Сосудистый цилиндр проходит через центр корня в области созревания и области удлинения. В области клеточного деления клетки значительно меньше. Клетки в этой области называются апикальной меристемой.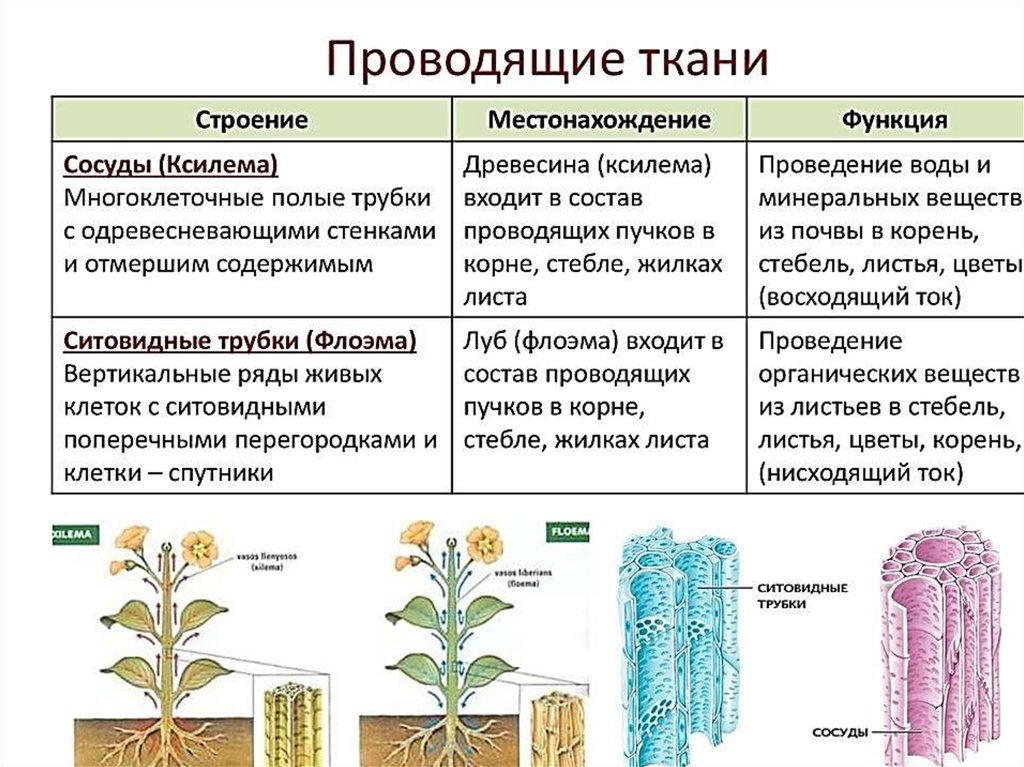 Апикальную меристему окружает слой клеток, называемый корневым чехликом. Изображение предоставлено: OpenStax Biology
Апикальную меристему окружает слой клеток, называемый корневым чехликом. Изображение предоставлено: OpenStax Biology
Растения также могут иметь боковые корни , которые ответвляются от основного стержневого корня. Боковые корни происходят из меристематической ткани перицикла , который является самым наружным клеточным слоем в сосудистом цилиндре в центре корня (показан ниже). После того, как они появились, боковые корни демонстрируют свой собственный первичный рост, постоянно увеличивая длину бокового корня.
Окрашивание показывает различные типы клеток на этой световой микрофотографии пшеницы ( Triticum ) поперечное сечение корня. Клетки склеренхимы экзодермы и клетки ксилемы окрашиваются в красный цвет, а клетки флоэмы — в синий. Другие типы клеток окрашиваются в черный цвет. Стела, или сосудистая ткань, представляет собой область внутри эндодермы (обозначена зеленым кольцом). Корневые волоски видны вне эпидермиса. (кредит: OpenStax Biology, данные масштабной линейки от Мэтта Рассела)
Изображение представляет собой поперечный срез части корня однодольного кукурузы ( Zea mays ), показывающий стелу и боковой корень. Боковые корни развиваются из слоя клеток под эндодермой, называемого перициклом. Они возникают путем клеточного деления клеток перицикла напротив группы протоксилем. Корень начинает набухать по мере того, как новый боковой корень проникает наружу к поверхности, пробивая себе путь через клетки кортикальной паренхимы и, наконец, прорываясь через эпидермис в почву. По мере роста в нем развиваются ксилема и флоэма, которые соединяются с сосудистыми тканями главного корня. Этот процесс сильно отличается от того, что происходит в побегах, где боковые ветви, образующие стебли или листья, происходят только из верхушечной меристематической ткани побега. Боковые корни могут развиваться на большом расстоянии от кончика корня. Изображение Джона Беббингтона FRPS, https://www.flickr.com/photos/71183136@N08/6947183226
Боковые корни развиваются из слоя клеток под эндодермой, называемого перициклом. Они возникают путем клеточного деления клеток перицикла напротив группы протоксилем. Корень начинает набухать по мере того, как новый боковой корень проникает наружу к поверхности, пробивая себе путь через клетки кортикальной паренхимы и, наконец, прорываясь через эпидермис в почву. По мере роста в нем развиваются ксилема и флоэма, которые соединяются с сосудистыми тканями главного корня. Этот процесс сильно отличается от того, что происходит в побегах, где боковые ветви, образующие стебли или листья, происходят только из верхушечной меристематической ткани побега. Боковые корни могут развиваться на большом расстоянии от кончика корня. Изображение Джона Беббингтона FRPS, https://www.flickr.com/photos/71183136@N08/6947183226
Первичный рост в побегах
Приведенная ниже информация была адаптирована из OpenStax Biology 30.2
Как и в корнях, первичный рост в стеблях является результатом быстрого деления клеток в апикальных меристемах на верхушке побега. Затем последующее удлинение клеток приводит к первичному росту.
Затем последующее удлинение клеток приводит к первичному росту.
У многих растений основной первичный рост происходит преимущественно в верхушечной (верхней) почке, а не в пазушных почках (почках в местах бокового ветвления). Влияние верхушечной почки на общий рост растений известно как верхушечное доминирование , препятствующее росту пазушных почек, образующихся по бокам ветвей и стеблей. Большинство хвойных деревьев демонстрируют сильное преобладание вершины, что приводит к типичной конической форме рождественской елки. Если верхушечную почку удалить, то пазушные почки начнут формировать боковые ответвления. Садоводы используют этот факт, когда обрезают растения, срезая верхушки ветвей, тем самым стимулируя рост пазушных почек, придавая растению кустистую форму.
Вторичный рост побегов (и корней)
Процесс вторичного роста контролируется боковыми меристемами и одинаков как для стеблей, так и для корней. Боковые меристемы включают сосудистый камбий, а у древесных растений — пробковый камбий (камбий — это еще один термин для меристемы). Травянистые (недревесные) растения в основном подвергаются первичному росту, почти без вторичного роста или увеличения толщины. У древесных растений заметен вторичный рост, или древесина; это встречается у некоторых двудольных, но очень редко у однодольных.
Травянистые (недревесные) растения в основном подвергаются первичному росту, почти без вторичного роста или увеличения толщины. У древесных растений заметен вторичный рост, или древесина; это встречается у некоторых двудольных, но очень редко у однодольных.
Приведенная ниже информация относится к вторичному росту стеблей. Хотя принципы вторичного роста корней аналогичны, детали несколько отличаются. Мы обсудим только детали, характерные для стеблей.
Сосудистый камбий расположен между первичной ксилемой и первичной флоэмой внутри сосудистого пучка. (Напомним, что ксилема расположена внутри, а флоэма — снаружи пучка.) Клетки сосудистого камбия делятся и образуют вторичную ксилему (трахеиды и элементы сосудов) внутри и вторичную флоэму (ситовидные элементы и клетки-спутники). наружу. Клетки вторичной ксилемы содержат лигнин , основной компонент древесины, обеспечивающий морозостойкость и прочность. Ксилема вместе с сердцевиной образуют древесину одеревеневшего стебля.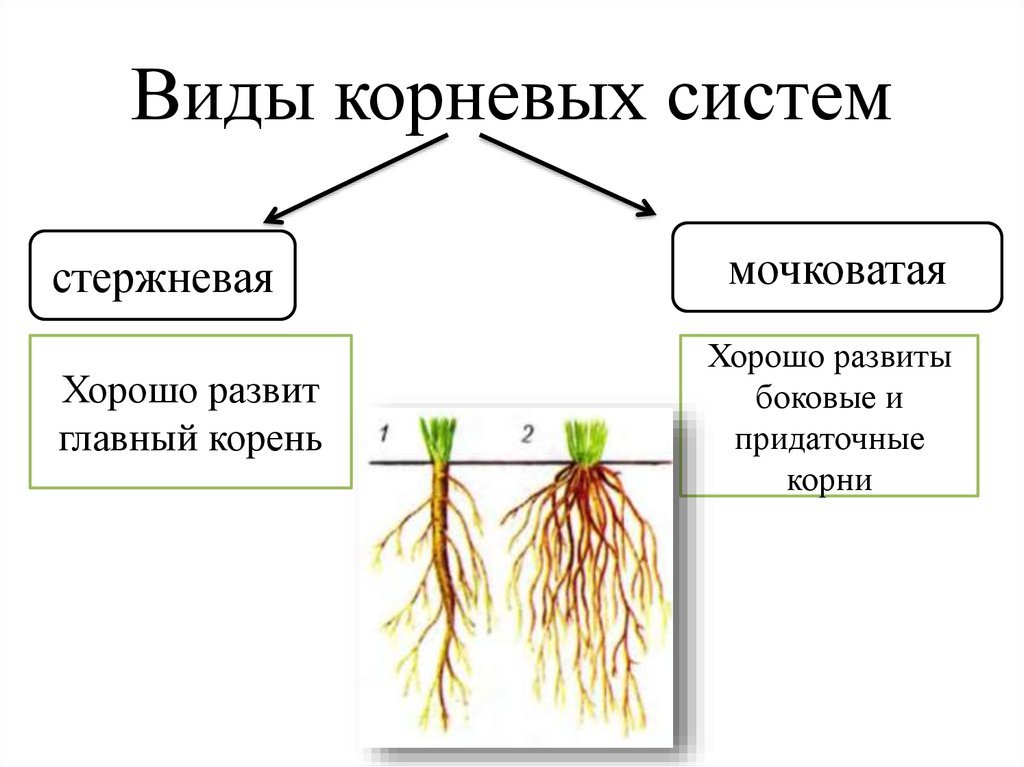
У древесных растений пробковый камбий является наружной боковой меристемой. Он производит пробковых клеток , которые содержат воскообразное вещество, способное отталкивать воду. Флоэма вместе с пробковыми клетками образует кору , которая защищает растение от физических повреждений и способствует уменьшению потери воды. Пробковый камбий также образует слой клеток, известный как 9.0123 феллодерма , растущая внутрь от камбия. Пробковый камбий, пробковые клетки и феллодерма вместе называются перидермой . Перидерма заменяет эпидермис у взрослых растений.
Совместное действие сосудистой и пробковой камбии приводит к вторичному росту или расширению стебля растения. Эти структуры показаны ниже:
У древесных растений за первичным ростом следует вторичный рост, который позволяет стеблю растения увеличиваться в толщине или в обхвате. Вторичная сосудистая ткань добавляется по мере роста растения, а также пробковый слой. Кора дерева простирается от сосудистого камбия до эпидермиса. Изображение предоставлено: OpenStax Biology
Кора дерева простирается от сосудистого камбия до эпидермиса. Изображение предоставлено: OpenStax Biology
Каждый год в течение вегетационного периода добавляется новый слой ксилемы и флоэмы. Внутренние слои ксилемы в конечном итоге отмирают и заполняются смолой, функционируя только в качестве структурной поддержки. Внутренняя, нефункциональная ксилема называется сердцевиной . Более новая функциональная ксилема называется заболонь . Наружные слои флоэмы со временем придавливаются пробковым камбием и разрушаются. Таким образом, зрелое дерево содержит много внутренних слоев старой, нефункциональной ксилемы глубоко внутри ствола, но лишь небольшое количество старой флоэмы.
Слои тканей ствола взрослого дерева. Изображение основано на работе Брата Лаппина — общественное достояние, https://commons.wikimedia.org/w/index.php?curid=8979988
В этом видео описывается процесс и результат вторичного роста стеблей:
Вторичные приросты и годовые кольца
Деятельность сосудистого камбия приводит к образованию годичных колец роста. В весенний вегетационный период клетки вторичной ксилемы имеют большой внутренний диаметр, а их первичные клеточные стенки не сильно утолщены. Это известно как ранняя древесина или весенняя древесина. В осенний сезон вторичная ксилема развивает утолщенные клеточные стенки, образуя позднюю или осеннюю древесину, которая более плотная, чем ранняя древесина. Такое чередование ранней и поздней древесины обусловлено в основном сезонным уменьшением числа сосудистых элементов и сезонным увеличением числа трахеид. Это приводит к образованию годового кольца, которое можно увидеть как круглое кольцо в поперечном сечении стебля (показано ниже).
Leave A Comment