
Когда появился фагоцитоз? • Сергей Ястребов • Новости науки на «Элементах» • Эволюция, Палеонтология, Микробиология
Фагоцитоз — очень важный тип клеточной активности, позволяющий клеткам целиком поглощать друг друга. Из всех живых организмов Земли фагоцитозом (насколько сейчас известно) обладают только эукариоты. Надежные свидетельства существования фагоцитоза появляются в палеонтологической летописи около миллиарда лет назад. Между тем ближайший общий предок всех современных эукариот (last eukaryotic common ancestor, LECA), судя по оценкам «молекулярных часов», жил примерно 1,6–1,8 миллиарда лет назад. Либо эукариоты приобрели фагоцитоз не сразу, либо первое время он не оставлял в палеонтологической летописи следов. Проверить это можно палеонтологическими находками: например, красные водоросли могли сформироваться только с помощью фагоцитоза, поэтому любая ископаемая находка красной водоросли означает, что к этому моменту истории Земли фагоцитоз уже точно был. Находки красных водорослей возрастом более миллиарда лет пока проблематичны. Есть и другие нерешенные вопросы. В любом случае на слишком раннее появление фагоцитоза (до LECA) пока ничто не указывает.
Есть и другие нерешенные вопросы. В любом случае на слишком раннее появление фагоцитоза (до LECA) пока ничто не указывает.
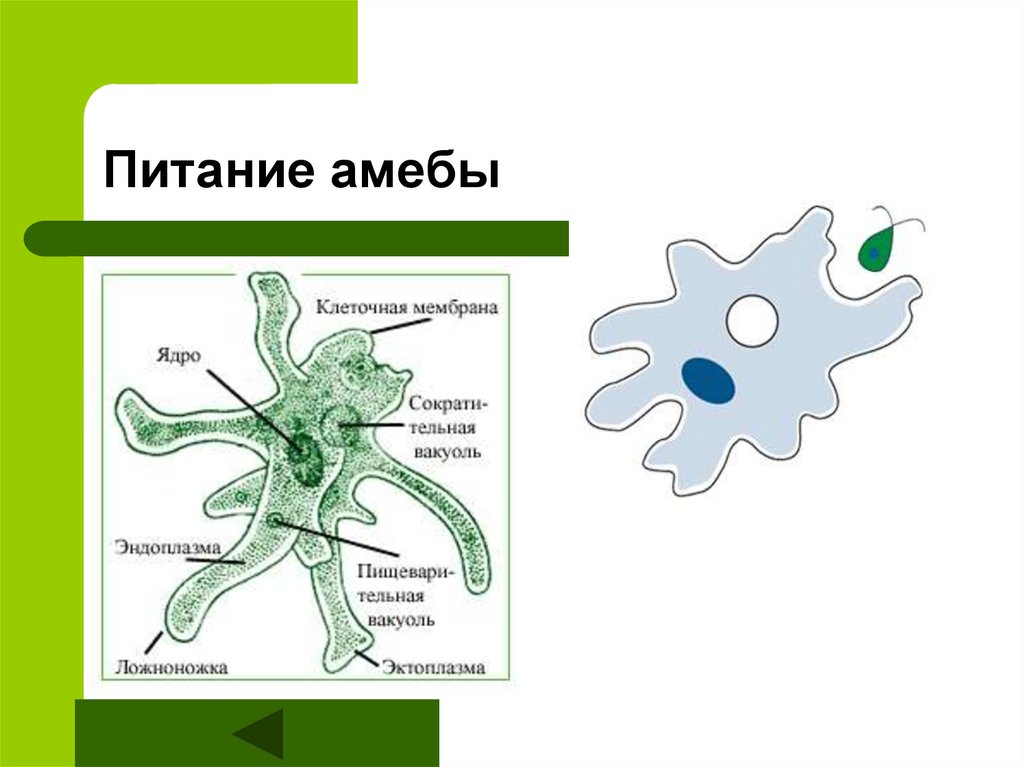
Фагоцитоз — очень важное свойство эукариотных клеток, которое открыл в конце XIX века великий русский биолог Илья Ильич Мечников. Работая с морской фауной, он обнаружил, что в телах губок, личинок морских звезд и других животных встречаются подвижные клетки, которые самостоятельно ползают, активно захватывают твердые частицы, а потом переваривают их у себя внутри (внутриклеточное пищеварение). Такие клетки были названы фагоцитами, то есть «пожирающими клетками», а само явление — фагоцитозом.
В современной научной литературе фагоцитоз определяют как активное (с помощью ложноножек) поглощение клеткой крупных частиц. «Крупные» в данном случае значит «видимые в световой микроскоп», то есть размером хотя бы в половину микрона. Посредством фагоцитоза эукариотная клетка может питаться бактериями, а может и другими эукариотами: так тоже бывает очень часто. Мир эукариот — это мир хищников и жертв.
Как известно, все живые организмы, состоящие из клеток, делятся на эукариот, бактерий и архей. Фагоцитоз — уникальное «ноу-хау» эукариот. Никакие известные бактерии или археи им не владеют. Клетка, способная к фагоцитозу, должна быть крупнее своей добычи (это очевидно) и должна иметь в своей цитоплазме сложную систему сократимых элементов, которые позволят ей быстро менять форму (см. Цитоскелет). Долгое время все думали, что белки, из которых можно построить такую систему, есть только у эукариот. В последние годы выяснилось, что это неверно: довольно большой набор белков, обычно связанных с внутриклеточной сократительной системой, есть у асгардархей (см. Asgardarchaeota) — группы архей, от которой эукариоты, по всей видимости, произошли (см. Описан новый надтип архей, к которому относятся предки эукариот, «Элементы», 16.01.2017). Однако надо учитывать, что это «вычислено» по биоинформатическим данным — по анализу последовательностей ДНК, найденных в донных грунтах и тому подобных местах.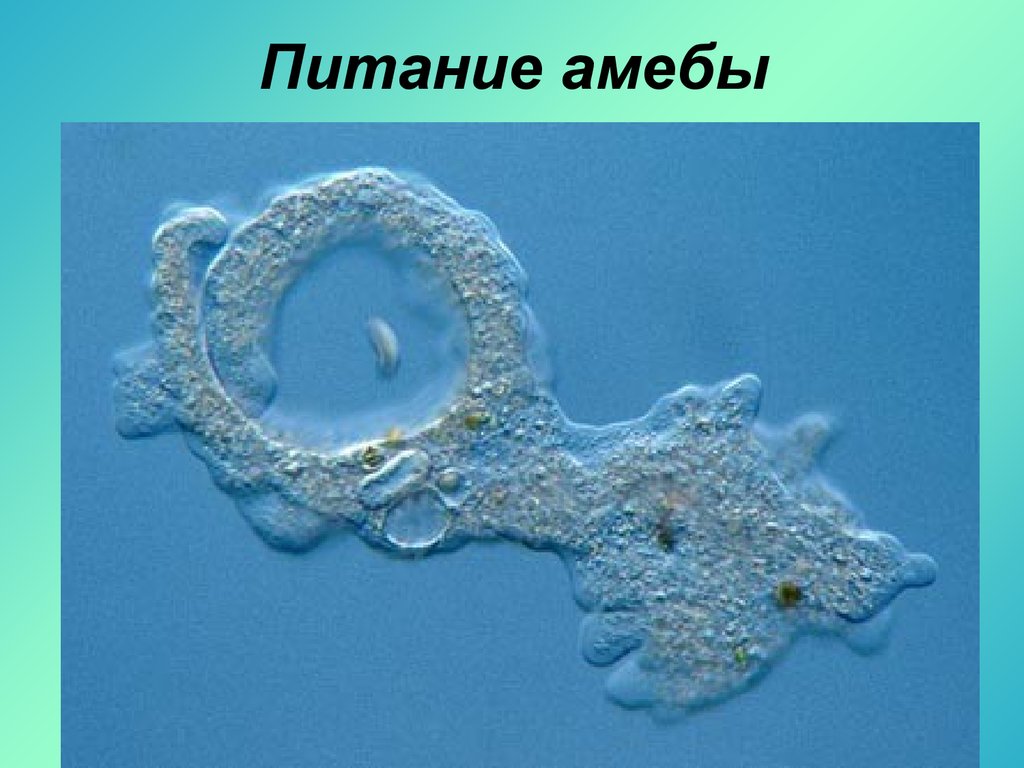
Более того, есть неплохо обоснованное мнение, что фагоцитоз просто-напросто не мог возникнуть раньше, чем сложился полный комплекс особенностей эукариотной клетки, включая ядро и митохондрии. Дело в том, что акт фагоцитоза сам по себе требует довольно больших вложений энергии: поймать, проглотить и переварить крупную добычу не так уж легко. Расчеты показывают, что без внутриклеточных «энергетических станций», которыми служат митохондрии, фагоцитоз как способ питания был бы убыточным: протоэукариотная клетка тратила бы больше энергии, чем получала (W. F. Martin et al., 2017. The physiology of phagocytosis in the context of mitochondrial origin). Так что версия, согласно которой фагоцитоз появился только у полноценных эукариот, по-прежнему заслуживает внимания.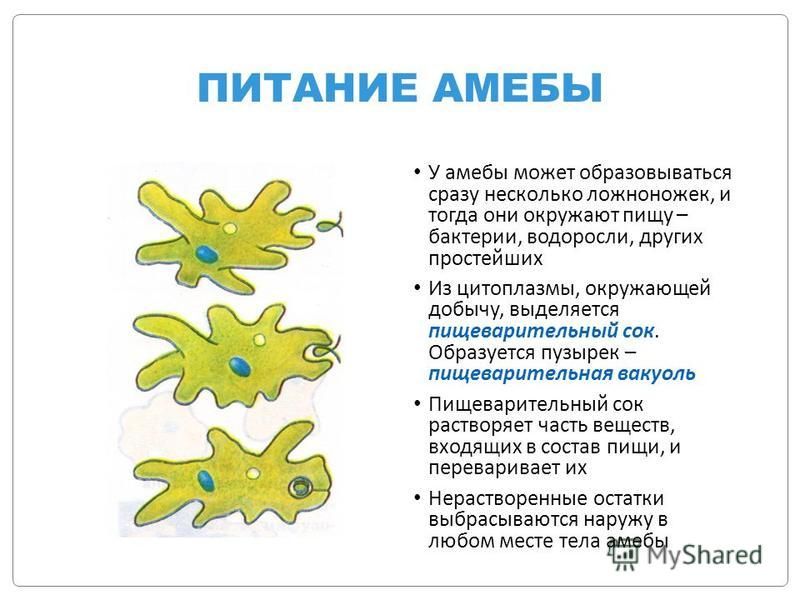
Когда же именно он появился? Для понимания истории жизни на Земле этот вопрос очень важен. Появление фагоцитоза, безусловно, стало для земных экосистем революционным событием. До «изобретения» фагоцитоза на Земле не было никаких хищников, способных целиком заглатывать другие живые организмы. Появление таких хищников — пусть даже одноклеточных — сразу изменило структуру пищевых цепочек, сделав их более длинными и сложными. Это не могло не привести к глобальным экологическим переменам. Кроме того, известно, что давление хищников способно «работать» своего рода ускорителем эволюции, стимулирующим появление новых форм за счет того, что жертва вынуждена приспосабливаться к хищнику (А. С. Раутиан, А. Г. Сенников, 2001. Отношения хищник-жертва в филогенетическом масштабе времени). Но и хищник, в свою очередь, приспосабливается к жертве. Начинается коэволюция, которая может увести и хищников, и жертв очень далеко. В конце концов, такое важнейшее эволюционное событие, как возникновение многоклеточных животных, было бы без фагоцитоза просто немыслимо.
Однако датировать появление фагоцитоза очень трудно. Первый обладатель фагоцитоза наверняка был бесскелетным одноклеточным существом, похожим на жгутиконосца или амёбу. Шансов сохраниться в палеонтологической летописи у него практически не было. Во всяком случае, надеяться на эти шансы не стоит. Лучше обратиться к анализу косвенных данных — здесь, к счастью, палеонтология может хоть что-то предложить.
Прежде всего, появление фагоцитоза имело зримые последствия в виде приобретения некоторыми эукариотами хлоропластов. Хлоропласты — это потомки фотосинтезирующих цианобактерий, которые были когда-то поглощены эукариотной клеткой, но не переварены, а «обращены в рабство»: они продолжают заниматься фотосинтезом, поставляя хозяину полезные продукты. Краткий обзор эволюционной истории хлоропластов можно найти в статье Открыт хищный родственник красных водорослей, «Элементы», 30.07.2019.
Только благодаря хлоропластам на свете смогли появиться фотосинтезирующие эукариоты — в том числе и многоклеточные, которых мы называем нестрогими терминами «водоросли» или «растения».
Самые древние обладатели хлоропластов относятся к супергруппе эукариот, которая называется Archaeplastida (см. Случайно открытый жгутиконосец обновляет систему эукариот, «Элементы», 06.02.2019). Это красные водоросли, глаукофитовые водоросли и зеленые растения. Хлоропласты архепластид — первичные: это означает, что они образовались непосредственно из захваченных цианобактерий (а не из других эукариот). Если в некоторый момент истории Земли мы видим фотосинтезирующих архепластид, значит, к этому моменту уже совершенно точно существовал фагоцитоз, позволяющий «глотать» как минимум бактерий. Здесь палеонтология может провести четкую верхнюю границу.
Итак, самый древний бесспорный представитель архепластид — это многоклеточная красная водоросль Bangiomorpha pubescens (рис. 2). Ее возраст, по последним данным, равен примерно 1,05 миллиарда лет (T. M. Gibson et al., 2018. Precise age of Bangiomorpha pubescens dates the origin of eukaryotic photosynthesis). При этом бангиоморфа поразительно похожа на современные бангиевые водоросли. За миллиард лет они умудрились почти не измениться. Это, пожалуй, самый выдающийся пример живых ископаемых среди всех эукариот. Современные бангиевые водоросли во много раз превосходят по древности, например, латимерию или гинкго, и заслуживают уж никак не меньшего почета. Жаль, что они относительно малоизвестны.
При этом бангиоморфа поразительно похожа на современные бангиевые водоросли. За миллиард лет они умудрились почти не измениться. Это, пожалуй, самый выдающийся пример живых ископаемых среди всех эукариот. Современные бангиевые водоросли во много раз превосходят по древности, например, латимерию или гинкго, и заслуживают уж никак не меньшего почета. Жаль, что они относительно малоизвестны.
Кроме того, описаны остатки многоклеточных водорослей — возможно, тоже красных — имеющие более древний возраст, а именно 1,6 миллиарда лет (S. Bengtson et al., 2016. Three-dimensional preservation of cellular and subcellular structures suggests 1.6 billion-year-old crown-group red algae). В отличие от бангиоморфы, с которой ситуация выглядит ясной, здесь авторы — среди них известный шведский палеонтолог Стефан Бенгтсон (Stefan Bengtson) — лишь предполагают принадлежность объектов к красным водорослям, основываясь на сохранившихся деталях структуры клеток. Но ничего невероятного в таком предположении нет.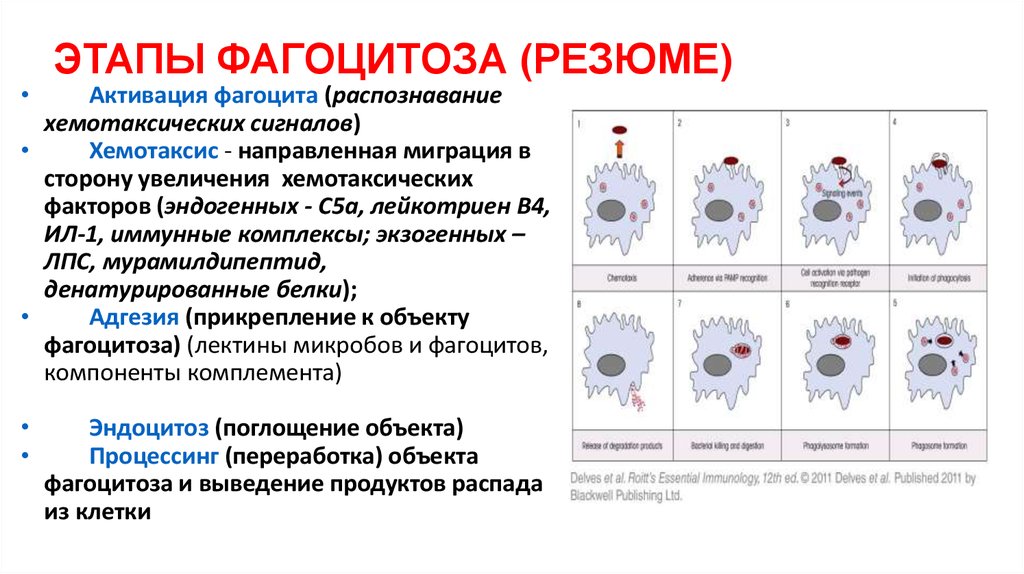
В земных слоях возрастом от 1,15 до 0,9 миллиарда лет назад начинают обнаруживаться остатки эукариотных водорослей, клеточные стенки которых пронизаны характерными перфорациями — проще говоря, дырками (рис. 3, C. C. Loron et al., 2018. Implications of selective predation on the macroevolution of eukaryotes: evidence from Arctic Canada). Такие перфорации проделывают в оболочках клеток своих жертв современные амёбы из группы вампиреллид (Vampyrellidae; см. также красочное описание в блоге
 Находка съеденных ими водорослей возрастом миллиард лет — первое прямое свидетельство существования эукариот, питающихся другими эукариотами.
Находка съеденных ими водорослей возрастом миллиард лет — первое прямое свидетельство существования эукариот, питающихся другими эукариотами.Наконец, около 760 миллионов лет назад в палеонтологической летописи появляются панцири раковинных амёб, относящихся к группе арцеллинид (S. M. Porter, L. A. Riedman, 2019. Evolution: ancient fossilized amoebae find their home in the tree). Надо сказать, что на вампиреллид эти амёбы совсем не похожи и даже им не родственны (они относятся к другой супергруппе). Арцеллиниды процветают до сих пор, в изобилии населяя пресные воду и почву и питаясь как бактериями, так и другими эукариотами (рис. 1). Из принципа актуализма следует, что они скорее всего вели примерно такой же образ жизни и 700 с лишним миллионов лет назад. Ну, а более молодые свидетельства фагоцитоза уже бессмысленно специально искать: они будут попадаться на каждом шагу.
Впечатления от всего этого получаются двоякие. Мы наглядно видим, насколько мощными бывают в естественных науках косвенные доводы. У красных водорослей фагоцитоза нет. Но если где-то в палеонтологической летописи мы обнаруживаем хотя бы одну красную водоросль, это с полной логической неизбежностью означает, что к данному историческому моменту фагоцитоз на Земле уже был. Никаким другим способом красные водоросли — а равно и любые другие эукариотные водоросли — просто не могли возникнуть (см. Открыт хищный родственник красных водорослей, «Элементы», 30.07.2019). В то же время перечисленные палеонтологические данные ничего не сообщают о ранних этапах эволюции эукариот. Времена, когда эукариоты только формировались, остаются «за кадром».
У красных водорослей фагоцитоза нет. Но если где-то в палеонтологической летописи мы обнаруживаем хотя бы одну красную водоросль, это с полной логической неизбежностью означает, что к данному историческому моменту фагоцитоз на Земле уже был. Никаким другим способом красные водоросли — а равно и любые другие эукариотные водоросли — просто не могли возникнуть (см. Открыт хищный родственник красных водорослей, «Элементы», 30.07.2019). В то же время перечисленные палеонтологические данные ничего не сообщают о ранних этапах эволюции эукариот. Времена, когда эукариоты только формировались, остаются «за кадром».
Можно зайти с другой стороны: внимательно посмотреть, что говорит о древности эукариот молекулярная филогенетика, а потом попытаться согласовать эти данные с палеонтологическими. Тут, однако, надо начать с важных оговорок. Современная биология работает с монофилетическими группами: любая такая группа происходит от одного общего предка и включает абсолютно всех его потомков. Корректно выделенная монофилетическая группа называется таксоном. Но общий предок любого крупного таксона жил много миллионов лет назад. А эволюция беспощадна: большая часть идущих от этого предка эволюционных линий наверняка давно вымерла. До современности дотянулись только те ветви, которым очень повезло. Вот их-то принято объединять в так называемую кроновую группу (crown group), которая начинается с общего предка всех современных членов данного таксона и включает всех его потомков. Остальная часть эволюционного древа таксона называется стволовой группой (stem group; см. Древнейшие предки кольчатых червей могли быть похожи на брахиопод, «Элементы», 26.02.2016). Все члены стволовых групп — по определению ископаемые.
Но общий предок любого крупного таксона жил много миллионов лет назад. А эволюция беспощадна: большая часть идущих от этого предка эволюционных линий наверняка давно вымерла. До современности дотянулись только те ветви, которым очень повезло. Вот их-то принято объединять в так называемую кроновую группу (crown group), которая начинается с общего предка всех современных членов данного таксона и включает всех его потомков. Остальная часть эволюционного древа таксона называется стволовой группой (stem group; см. Древнейшие предки кольчатых червей могли быть похожи на брахиопод, «Элементы», 26.02.2016). Все члены стволовых групп — по определению ископаемые.
В случае с эукариотами молекулярные данные, разумеется, доступны только для членов кроновой группы. Только для них и можно построить молекулярное филогенетическое древо. Члены стволовой группы эукариот жили сотни миллионов (если не миллиарды) лет назад, и никакой ДНК от них не осталось. Таким образом, фигурирующий в научных статьях последний эукариотный общий предок (last eukaryotic common ancestor, LECA), — это общий предок именно кроновой группы (см. Crown eukaryotes).
Crown eukaryotes).
Но все же древо кроновой группы — это гораздо лучше, чем ничего. Тут, правда, возникает еще одна трудность: датировки всевозможных эволюционных событий по молекулярным часам становятся надежными, только если эти «часы» калибруются по палеонтологическим данным (см. Палеонтологи уточнили время появления первых членистоногих, «Элементы», 23.08.2018), а для древнейших эукариот таких данных либо нет, либо они очень фрагментарны. Поэтому неудивительно, что в датировках имеется разброс. Но почти все они вмещаются в интервал между одним и двумя миллиардами лет назад. Типичная молекулярная оценка времени жизни LECA: 1,77 миллиарда лет назад плюс-минус несколько десятков миллионов лет (L. W. Parfrey et al., 2011. Estimating the timing of early eukaryotic diversification with multigene molecular clocks).
Допустим, что эта датировка верна. Но тогда мы сразу обнаруживаем, что последний общий предок всех современных эукариот (LECA) жил почти на 800 миллионов лет раньше, чем в летописи появляются надежные свидетельства эукариотного хищничества, про которые мы говорили выше.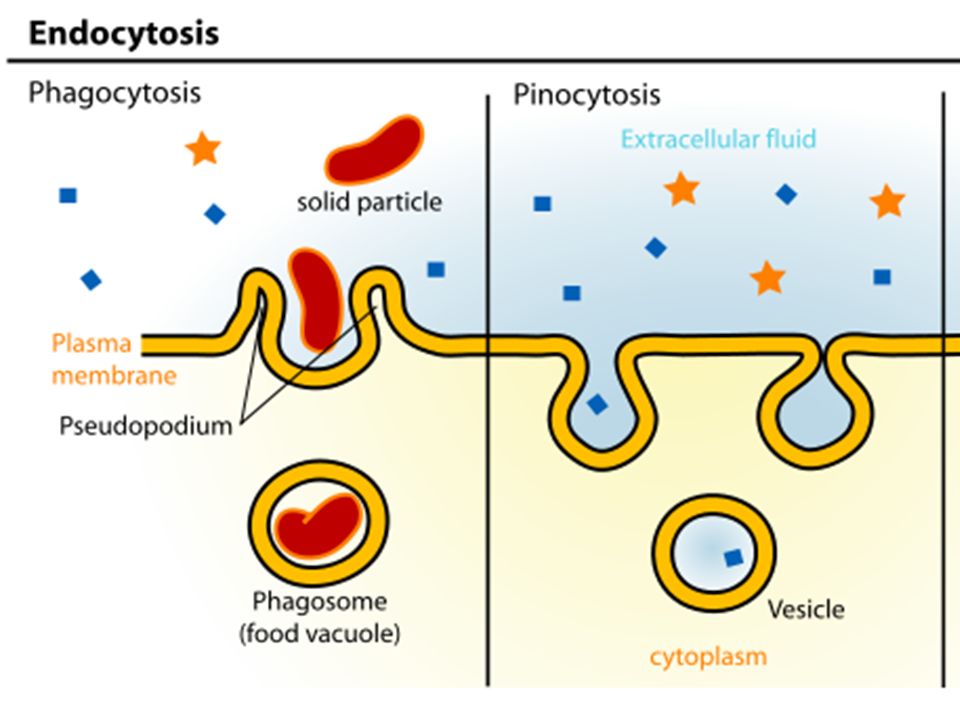 И как же это объяснить?
И как же это объяснить?
Что ж, объяснений можно предложить как минимум три.
Первое: LECA не был хищником. Он не владел фагоцитозом, а питался осмотрофно, всасывая растворенные вещества через клеточную мембрану. Фагоцитоз появился на несколько сот миллионов лет позже, и скорее всего — независимо в разных эволюционных ветвях (они безусловно успели к тому времени разойтись).
Второе: выбранная нами датировка неверна, LECA жил гораздо позднее. Есть и такие работы, где его наиболее вероятный возраст определяется всего лишь в 1,1 миллиарда лет (C. Berney, J. Pawlowski, 2006. A molecular time-scale for eukaryote evolution recalibrated with the continuous microfossil record). Тогда разрыв между LECA и «выходом на сцену» эукариотных хищников исчезает: ничто не противоречит тому, что LECA и был первым из этих хищников.
Третье: компромиссный вариант, LECA имеет скорее более древний, чем более молодой возраст (не менее 1,6 миллиарда лет; см. H. C. Betts et al., 2018. Integrated genomic and fossil evidence illuminates life’s early evolution and eukaryote origin), но фагоцитозом он все же обладал — просто ранние свидетельства активности хищных эукариот не сохранились или сохранились плохо.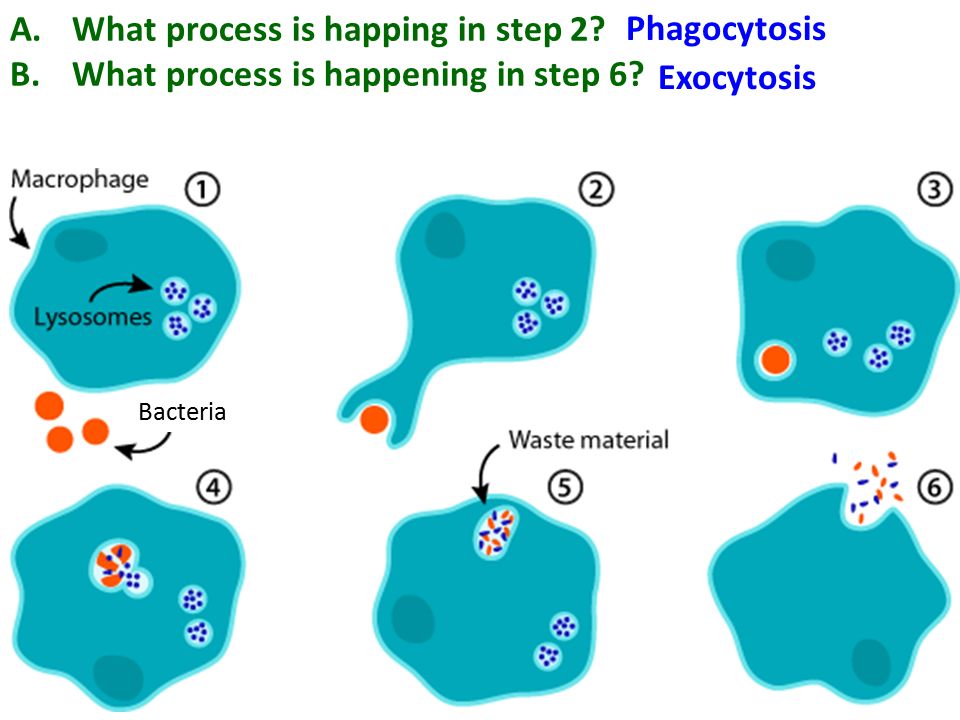
Первое объяснение противоречит широко принятому мнению, что LECA был жгутиконосцем, который питался бактериями. Правда, это мнение, пусть и распространенное, еще не обязательно должно быть верным. Есть и альтернативная гипотеза, согласно которой последний общий предок эукариот был осмотрофом, то есть «всасывальщиком» — примерно как грибы (W. Martin et al., 2003. Early cell evolution, eukaryotes, anoxia, sulfide, oxygen, fungi first (?), and a tree of genomes revisited). Но чем лучше мы узнаём древо эукариот, тем популярность этой гипотезы меньше. У самых примитивных современных одноклеточных эукариот — таких, как коллодиктиониды, малавимонады (Malawimonadidae) или анкиромонады (Ancyromonadida) — фагоцитоз вполне развит (см. Выделена новая супергруппа эукариот, «Элементы», 17.04.2018). В таких случаях обычно автоматически предполагают, что свойства, которые есть у нескольких ближайших потомков, были и у предка. Альтернатива должна означать, что фагоцитоз независимо возникал в разных ветвях эукариот буквально десятки раз, то есть мы имеем дело с мощнейшим параллелизмом.
Проблема второго объяснения в том, что оно слишком «омолаживает» LECA, выбирая в довольно широком спектре возможных датировок самую крайнюю точку. Такое непременно нуждается в дополнительных подтверждениях.
Слабость третьего объяснения в том, что оно опирается не столько на факты, сколько на их отсутствие. К сожалению, в палеонтологии так бывает часто. Впрочем, если данные о красных водорослях возрастом 1,6 миллиарда лет подтвердятся, то большой разрыв между эпохой LECA и появлением этих водорослей опять-таки исчезнет — и тогда вновь окажется, что LECA и был первым хищником. Так тоже может быть.
Ни одна из этих трех версий, однако, не предполагает, что фагоцитоз возник раньше LECA. Раннее (намного раньше LECA) возникновение фагоцитоза обосновать труднее всего. Оно не подтверждается ни классической палеонтологией (нет типичных водорослей, нет следов охоты эукариот друг на друга, нет защитных структур вроде тех же раковинок амеб-арцеллинид), ни палеоэкологией (нет подходящей по времени революции в сообществах, которую можно было бы связать с настолько ранним приходом хищников).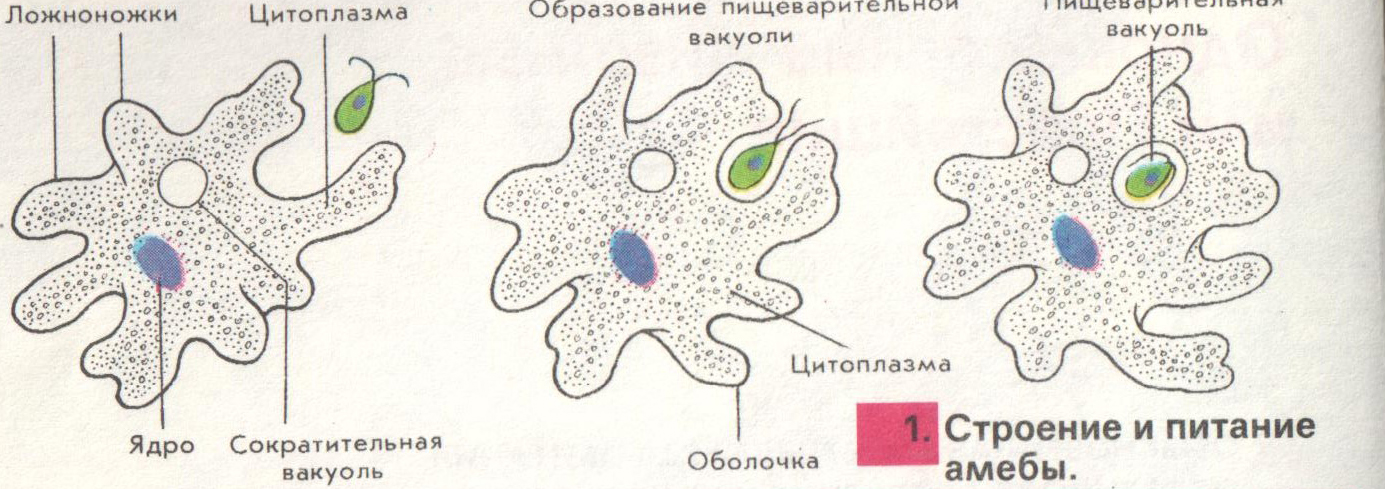 В этой связи надо вспомнить, что многочисленные гипотезы происхождения эукариот давно уже делятся на «фаготрофные», согласно которым предок эукариот просто «проглотил» предка митохондрий, и «синтрофные»: предок эукариотной клетки и предок митохондрий жили в мирном метаболическом симбиозе, не предполагавшем никакой охоты друг на друга (см. Обнаружен живой представитель асгардархей, «Элементы», 22.08.2019). Палеонтология на данный момент высказывается скорее за синтрофную версию, хотя здесь возникает очень много разных «если».
В этой связи надо вспомнить, что многочисленные гипотезы происхождения эукариот давно уже делятся на «фаготрофные», согласно которым предок эукариот просто «проглотил» предка митохондрий, и «синтрофные»: предок эукариотной клетки и предок митохондрий жили в мирном метаболическом симбиозе, не предполагавшем никакой охоты друг на друга (см. Обнаружен живой представитель асгардархей, «Элементы», 22.08.2019). Палеонтология на данный момент высказывается скорее за синтрофную версию, хотя здесь возникает очень много разных «если».
Например, совершенно непонятно, что делать с грибоподобными организмами, возраст остатков которых предположительно достигает 2,4 миллиарда лет (см. В вулканических породах возрастом 2,4 млрд лет найдены следы древнейших грибов, «Элементы», 11.05.2017). Грибы — это одна из множества ветвей типичных современных эукариот. Если они действительно имеют такую огромную древность, это опрокидывает все приведенные выше выкладки. Но пока есть сомнения как насчет их возраста, так и в том, что это вообще остатки живых существ (S. McMahon, 2019. Earth’s earliest and deepest purported fossils may be iron-mineralized chemical gardens). Тут палеонтологам надо еще разбираться.
McMahon, 2019. Earth’s earliest and deepest purported fossils may be iron-mineralized chemical gardens). Тут палеонтологам надо еще разбираться.
Источник: Danel B. Mills. The origin of phagocytosis in Earth history // Interface Focus. 2020. DOI: 10.1098/rsfs.2020.0019.
См. также:
Палеобиологи обнаружили древних амеб-вампиров, Lenta.ru, 26.05.2016.
Сергей Ястребов
Квалификационная работа магистра: «Изучение тонкого строения клетки и процесса фагоцитоза у амеб рода Paradermamoeba.» Аннотация: Покровы клетки голых лобозных амеб представлены плазматической мембраной, несущей слой гликокаликса, который может быть весьма сложно дифференцирован и достигать значительной толщины. Все взаимодействия клетки с окружающей средой – адгезия, фагоцитоз, пиноцитоз опосредованы гликокаликсом. Крупные пищевые объекты амебы поглощают с помощью фагоцитоза, в процессе которого клетка, образующая пищевую вакуоль, неизбежно заключает в нее часть собственного покрова, и вынуждена его переваривать, а потом восстанавливать на своей поверхности de novo. Направление: Биология Кафедра: Зоологии беспозвоночных Научный руководитель: к.б.н. Смирнов Алексей.Валерьевич Отзыв научного руководителя Рецензент: к. Рецензия Дата защиты: 9-06-2015 |
 Для амеб, обладающих особо толстым и высокодифференцированным покровом подобный процесс может оказаться весьма энергозатратным – существенная часть полученных в результате фагоцитоза ресурсов в этом случае уйдет на восстановление покрова, переваренного клеткой.
Для амеб, обладающих особо толстым и высокодифференцированным покровом подобный процесс может оказаться весьма энергозатратным – существенная часть полученных в результате фагоцитоза ресурсов в этом случае уйдет на восстановление покрова, переваренного клеткой. 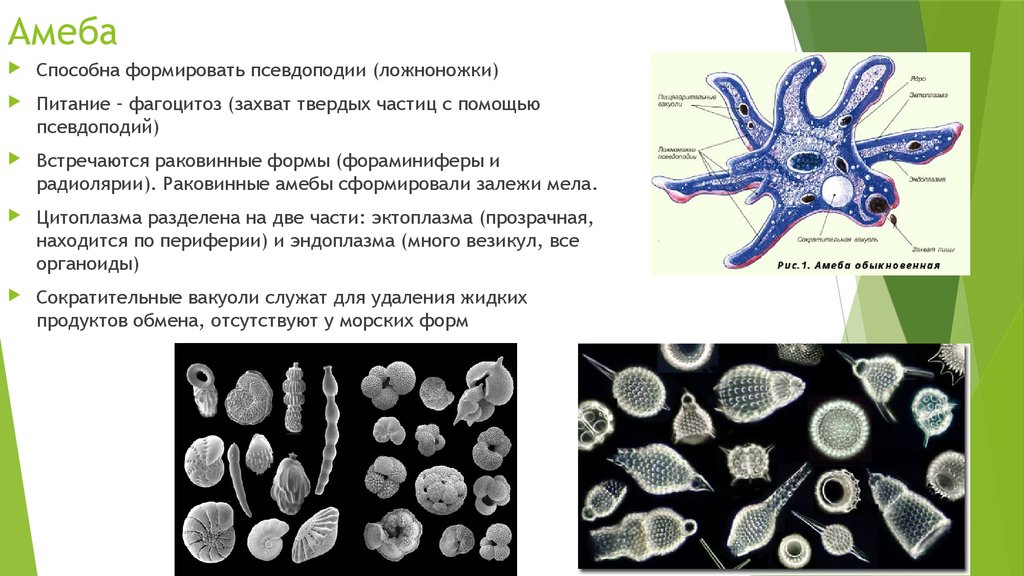 В клетках Paradermamoeba levis нами были впервые обнаружены цитоплазматические ЦОМТы ассоциированные с диктиосомами аппарата Гольджи. Обнаружение ЦОМТов позволяет пополнить список представителей подкласса Longamoebia, имеющих подобные структуры, и подтверждает, что их наличие, скорее всего, является синапоморфным признаком этой филогенетической ветви Amoebozoa. Нам удалось получить электронограммы процесса фагоцитоза у Paradermamoeba levis и впервые показать, что эти амебы исключают толстый дифференцированный гликокаликс из формирующейся пищевой вакуоли, подобно тому как это делают другие виды амеб, обладающие толстым высокодифференцированным поровом клетки. Таким образом, наши данные подтверждают гипотезу о неизбежном возникновении специфических способов питания у всех амеб с толстым высокодифференцированным гликокаликсом.
В клетках Paradermamoeba levis нами были впервые обнаружены цитоплазматические ЦОМТы ассоциированные с диктиосомами аппарата Гольджи. Обнаружение ЦОМТов позволяет пополнить список представителей подкласса Longamoebia, имеющих подобные структуры, и подтверждает, что их наличие, скорее всего, является синапоморфным признаком этой филогенетической ветви Amoebozoa. Нам удалось получить электронограммы процесса фагоцитоза у Paradermamoeba levis и впервые показать, что эти амебы исключают толстый дифференцированный гликокаликс из формирующейся пищевой вакуоли, подобно тому как это делают другие виды амеб, обладающие толстым высокодифференцированным поровом клетки. Таким образом, наши данные подтверждают гипотезу о неизбежном возникновении специфических способов питания у всех амеб с толстым высокодифференцированным гликокаликсом.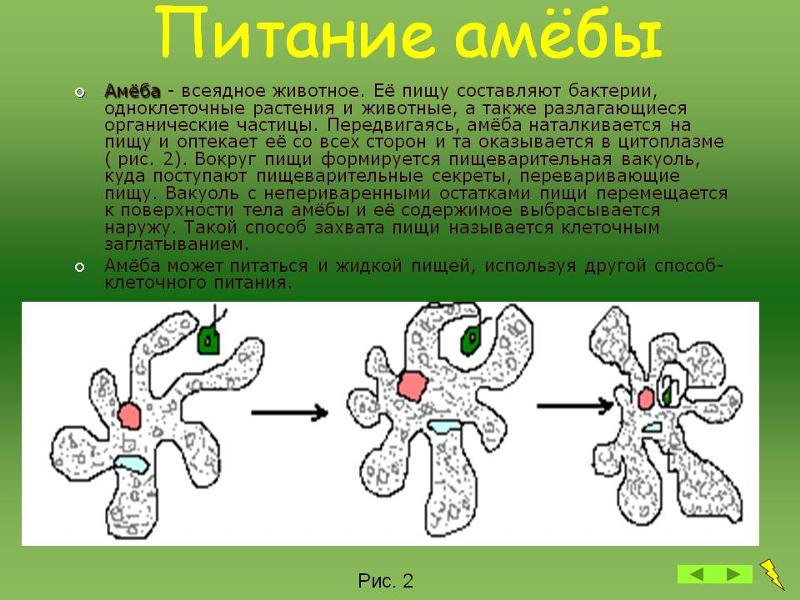 б.н. Чистякова Л.В.
б.н. Чистякова Л.В.Фагоцитоз | Определение, процесс и примеры
фагоцитоз
Просмотреть все СМИ
- Ключевые люди:
- Эли Мечников Карл Альберт Людвиг Ашофф
- Похожие темы:
- лейкоцит аутофагия макрофаг фагоцит ячейка Купфера
Просмотреть весь связанный контент →
фагоцитоз , процесс, при котором определенные живые клетки, называемые фагоцитами, поглощают или поглощают другие клетки или частицы. Фагоцит может быть свободноживущим одноклеточным организмом, таким как амеба, или одной из клеток организма, такой как лейкоцит. У некоторых форм жизни животных, таких как амебы и губки, фагоцитоз является средством питания. У высших животных фагоцитоз является главным образом защитной реакцией против инфекции и проникновения в организм чужеродных веществ (антигенов).
Ранние наблюдения
Присутствие инородных частиц внутри клеток впервые было описано в 1860-х годах патологом Кранидом Славянским. В 1880-х годах русский зоолог и микробиолог Эли Мечников ввел термин фагоцит для обозначения иммунных клеток, которые поглощают и уничтожают инородные тела, такие как бактерии. Мечников также признал, что фагоциты играют важную роль в иммунном ответе, открытие, которое принесло ему долю Нобелевской премии 1908 года по физиологии и медицине.
В 1880-х годах русский зоолог и микробиолог Эли Мечников ввел термин фагоцит для обозначения иммунных клеток, которые поглощают и уничтожают инородные тела, такие как бактерии. Мечников также признал, что фагоциты играют важную роль в иммунном ответе, открытие, которое принесло ему долю Нобелевской премии 1908 года по физиологии и медицине.
Типы фагоцитов
Наблюдайте фагоцитоз иммунной реакции человека, при котором лейкоциты поглощают или поглощают вторгшиеся микроскопические организмы или частицы
См. все видео к этой статье различные частицы пыли, пигменты и другие мельчайшие инородные тела. У человека и у позвоночных в целом наиболее эффективными фагоцитарными клетками являются два вида лейкоцитов: макрофаги (крупные фагоцитирующие клетки) и нейтрофилы (разновидность гранулоцитов). Макрофаги встречаются особенно в легких, печени, селезенке и лимфатических узлах, где их функция заключается в освобождении дыхательных путей, крови и лимфы от бактерий и других частиц. Макрофаги также обнаруживаются во всех тканях в виде блуждающих амебоидных клеток, а моноциты, предшественники макрофагов, обнаруживаются в крови. Меньшие фагоциты представляют собой в основном нейтрофилы, которые переносятся циркулирующей кровью до тех пор, пока не достигают области инфицированной ткани, где они проходят через стенку кровеносного сосуда и оседают в этой ткани. И макрофаги, и нейтрофилы притягиваются к области инфекции или воспаления с помощью веществ, выделяемых бактериями и инфицированной тканью, или посредством химического взаимодействия между бактериями и системой комплемента белков сыворотки крови. Нейтрофилы могут также поглощать частицы после случайного столкновения с ними.
Макрофаги также обнаруживаются во всех тканях в виде блуждающих амебоидных клеток, а моноциты, предшественники макрофагов, обнаруживаются в крови. Меньшие фагоциты представляют собой в основном нейтрофилы, которые переносятся циркулирующей кровью до тех пор, пока не достигают области инфицированной ткани, где они проходят через стенку кровеносного сосуда и оседают в этой ткани. И макрофаги, и нейтрофилы притягиваются к области инфекции или воспаления с помощью веществ, выделяемых бактериями и инфицированной тканью, или посредством химического взаимодействия между бактериями и системой комплемента белков сыворотки крови. Нейтрофилы могут также поглощать частицы после случайного столкновения с ними.Адгезия частиц
Изучить окрашенные изображения, позволяющие отличить фагоциты от бактерий во время фагоцитоза
Посмотреть все видео к этой статье До того, как фагоцитоз будет осуществлен, фагоцит и частица должны слипнуться друг с другом, возможность чего во многом зависит от химической природы поверхность частицы.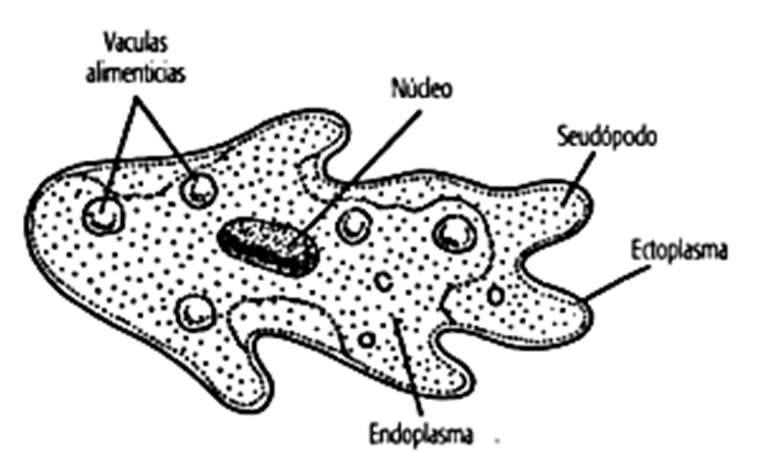 В случае бактерий, если фагоцит не может прикрепиться напрямую, белковые компоненты крови, известные как опсонины (например, комплемент и антитела), образуют поверхностную пленку на бактериях — процесс, известный как опсонизация. Фагоциты присоединяются к опсонинам, после чего следует фагоцитоз. Инкапсулированные бактерии попадают в организм с большим трудом. В отсутствие специфических антител, распознающих бактерии, опсонизация не может происходить, и бактерии отталкивают фагоциты. Поверхности таких бактерий покрываются специальными антителами только после того, как в организме вырабатывается иммунный ответ на присутствие данного вида бактерий. Такие антитела имеют большое значение в установлении иммунитета к заболеваниям.
В случае бактерий, если фагоцит не может прикрепиться напрямую, белковые компоненты крови, известные как опсонины (например, комплемент и антитела), образуют поверхностную пленку на бактериях — процесс, известный как опсонизация. Фагоциты присоединяются к опсонинам, после чего следует фагоцитоз. Инкапсулированные бактерии попадают в организм с большим трудом. В отсутствие специфических антител, распознающих бактерии, опсонизация не может происходить, и бактерии отталкивают фагоциты. Поверхности таких бактерий покрываются специальными антителами только после того, как в организме вырабатывается иммунный ответ на присутствие данного вида бактерий. Такие антитела имеют большое значение в установлении иммунитета к заболеваниям.
Поглощение и переваривание частиц
Скорость, с которой фагоцитирующая клетка поглощает частицу, зависит от размера частицы. Мелкие частицы, такие как бактерии или мельчайшие крупинки древесного угля, проглатываются почти мгновенно. Более крупные объекты, такие как скопления бактерий или клеток тканей, подвергаются фагоцитозу в ходе более продолжительного ответа. Клетка обтекает объект до тех пор, пока он не будет полностью поглощен. Таким образом, поглощенный объект заключен внутри мембраносвязанной вакуоли, называемой фагосомой. Фагоцит переваривает проглоченную частицу с помощью гидролитических ферментов, которые содержатся в окруженных мембраной мешочках, называемых лизосомами, которые находятся внутри клетки. Фагоцитарные ферменты секретируются в вакуоли, в которых происходит пищеварение. Мелкие органические компоненты частицы используются для построения более крупных молекул, необходимых клетке.
Клетка обтекает объект до тех пор, пока он не будет полностью поглощен. Таким образом, поглощенный объект заключен внутри мембраносвязанной вакуоли, называемой фагосомой. Фагоцит переваривает проглоченную частицу с помощью гидролитических ферментов, которые содержатся в окруженных мембраной мешочках, называемых лизосомами, которые находятся внутри клетки. Фагоцитарные ферменты секретируются в вакуоли, в которых происходит пищеварение. Мелкие органические компоненты частицы используются для построения более крупных молекул, необходимых клетке.
Эта статья была недавно отредактирована и обновлена Карой Роджерс.
Аутофагия | биология | Британика
аутофагия
Смотреть все СМИ
- Ключевые люди:
- Ёсинори Осуми
- Похожие темы:
- фагоцитоз лизосома
Просмотреть весь связанный контент →
аутофагия , также называемая аутофагоцитозом , деградация изношенных, аномальных или неисправных клеточных компонентов, которая происходит в органеллах, известных как лизосомы.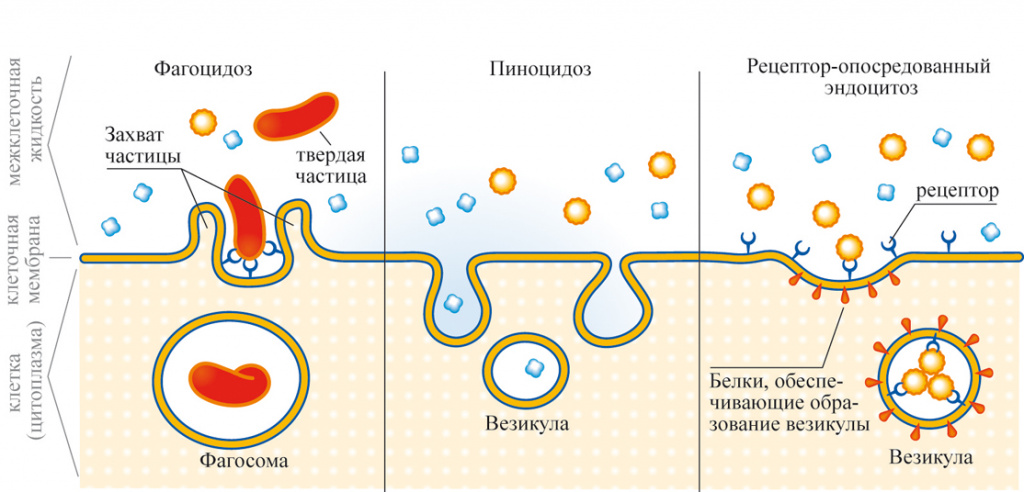 Аутофагия выполняет функции домашнего хозяйства, позволяя расщеплять и перерабатывать клеточные материалы, а также помогает сбалансировать потребности в энергии в периоды стресса. Термин аутофагия был введен в 1963 году бельгийским цитологом и биохимиком Кристианом Рене де Дювом, работа которого также предоставила первые доказательства участия лизосом в аутофагическом процессе.
Аутофагия выполняет функции домашнего хозяйства, позволяя расщеплять и перерабатывать клеточные материалы, а также помогает сбалансировать потребности в энергии в периоды стресса. Термин аутофагия был введен в 1963 году бельгийским цитологом и биохимиком Кристианом Рене де Дювом, работа которого также предоставила первые доказательства участия лизосом в аутофагическом процессе.
Известны три типа аутофагии: макроаутофагия, микроаутофагия и аутофагия, опосредованная шаперонами. Клетки в первую очередь полагаются на макроаутофагию, при которой изношенные или поврежденные клеточные материалы в цитозольных областях (заполненные жидкостью области, окружающие органеллы) клеток поглощаются аутофагосомами (везикулы с двойными мембранами, которые доставляют свое содержимое в лизосомы, где материалы деградируют). . При микроаутофагии клеточные компоненты поглощаются непосредственно через инвагинации лизосомальной мембраны. Опосредованная шаперонами аутофагия представляет собой избирательный процесс, при котором белок, известный как шаперон hsc70, распознает и связывается с белковыми субстратами, содержащими определенный аминокислотный мотив. Субстрат-мишень переносится в лизосому, где затем транслоцируется через мембрану посредством процесса, опосредованного рецептором.
Субстрат-мишень переносится в лизосому, где затем транслоцируется через мембрану посредством процесса, опосредованного рецептором.
Несколько генов кодируют различные компоненты механизма аутофагии, необходимого для секвестрации, транспорта, деградации и переработки клеточных материалов. Ферменты, кодируемые несколькими генами аутофагии, конъюгируют (соединяются вместе), тем самым повышая активность фермента, особенно во время образования аутофагосомы. Ряд неспецифических клеточных путей также имеет решающее значение для аутофагии, включая различные секреторные и эндоцитарные (поглощение) пути. Кроме того, цитоскелет, по-видимому, выполняет множественные функции в аутофагии, ключевую среди которых играет роль микротрубочек в облегчении транспорта аутофагосом в клетках млекопитающих.
В дополнение к своим функциям «домашнего хозяйства» и реакции на стресс, аутофагия также способствует иммунитету, помогая защищать клетки от болезнетворных организмов и участвуя в презентации антигена.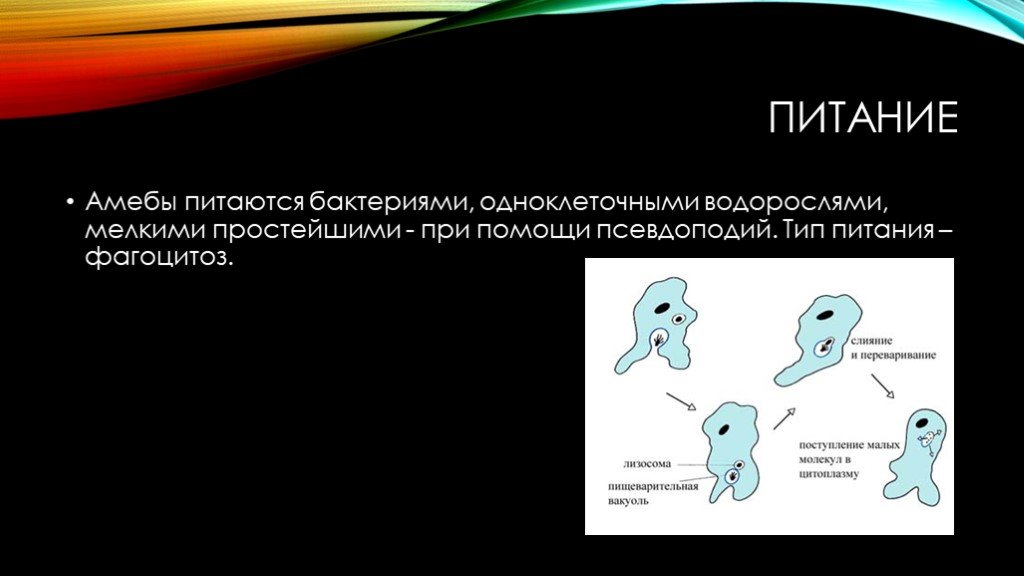
Leave A Comment