
черты и выявления приспособлений, относительный характер приспособительных признаков, механизм возникновения
Биология
12.11.21
10 мин.
Приспособленность организма к среде обитания играет огромное значение в процессе выживания живых существ и является результатом естественного отбора.
Оглавление:
- Приспособленность – что это
- Черты приспособленности к среде обитания и их результат
- Географическое видообразование
- Экологическое видообразование
- Относительный характер приспособленности кактуса
- Относительный характер приспособленности белого медведя
- Относительный характер приспособленности крота
- Относительный характер приспособленности верблюда
- Чем характеризуется приспособленность растений к опылению насекомыми
- Вывод о приспособленности организмов к среде обитания
Существование эволюционного механизма приспособленности обеспечивает максимальную адаптацию к условиям, в которых обитает вид.
Приспособленность – что это
Она заключается в соответствии особенностей строения, физиологических процессов и поведения живого организма той среде, в которой он обитает.
Этот механизм увеличивает шансы на выживание, оптимальное питание, спаривание и выращивание здорового потомства. Это универсальная особенность, свойственная для всех существ планеты от бактерий до высших форм жизни.
Данный механизм адаптации проявляется очень разнообразно. Растения, животные, рыбы, птицы, насекомые и прочие представители флоры и фауны довольно изобретательны в выборе средств, способствующих сохранению их вида.
Результатом становятся изменение окраски, формы тела, строения органов, способов размножения и питания.
Черты приспособленности к среде обитания и их результат
Например, тело лягушки сливается с цветом воды, травы и делает ее незаметной для хищников. Заяц-беляк меняет цвет с серого на белый зимой, что помогает ему быть незаметным на фоне снега.
Чемпионом в маскировочной практике считается хамелеон. Но, увы, мнение о том, что он подстраивается под цвет места, в котором находится, несколько упрощает реальную картину. Изменение цвета этой удивительной ящерицы является ответом на воздействие температуры воздуха, солнечных УФ лучей, и даже зависит от настроения.
Голова дятла – отличный пример формирования определенной формы тела, строения и функционирования органов. Птица обладает мощным, но упругим клювом, очень длинным тонким языком и системой амортизации, которая бережет мозг от травматизации при сильнейших ударах клюва птицы о ствол дерева.
Любопытная находка – “агрессия” у растений. Жгучие лепестки крапивы – отличный способ защиты от травоядных.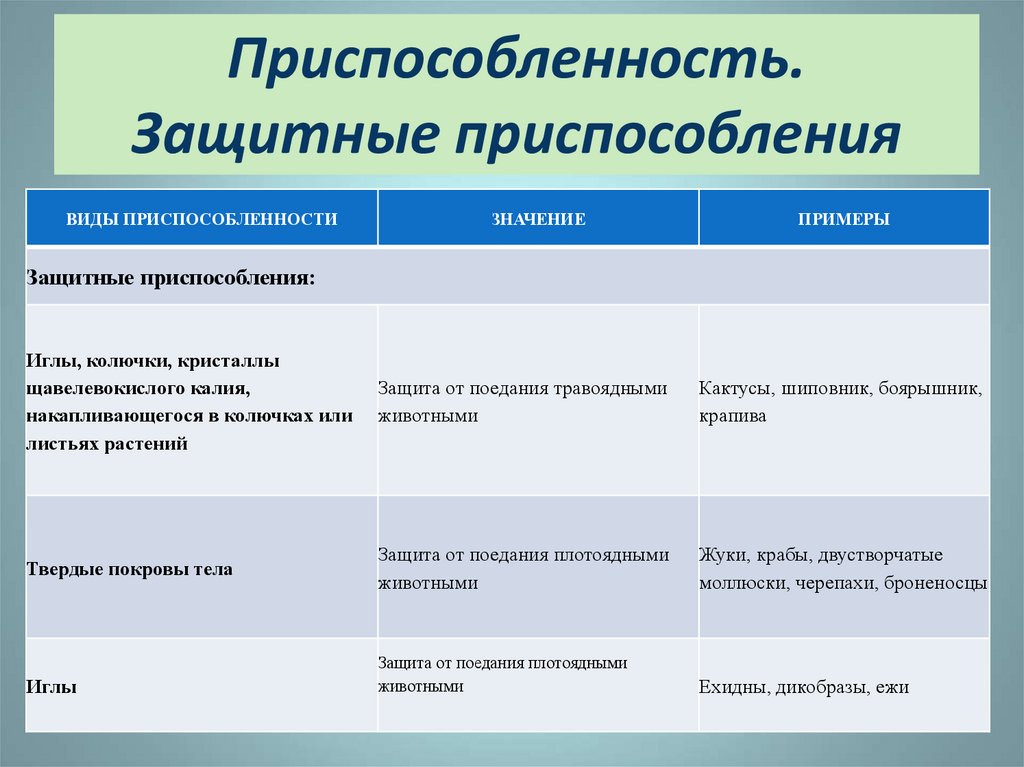
Географическое видообразование
Также уместно употребление термина “аллопатрическое” образование видов. Оно связано с расширением ареала обитания, когда вид занимает все большие территории. Или с тем, что территория разделяется естественными преградами – реками, горами и пр.
В такой ситуации происходит столкновение с новыми условиями и новыми “соседями” – видами, с которыми нужно учиться взаимодействию. Со временем это приводит к тому, что благодаря умению приспосабливаться у вида идет формирование и генетическое закрепление новых выгодных черт.
Представители географически изолированных популяций не скрещиваются между собой. В результате они начинают обладать рядом довольно ярких отличий от сородичей. Так, сумчатый волк и волк из отряда хищных в результате отбора довольно далеко разошлись по своим чертам.
Экологическое видообразование
Не связано с непосредственным расширением ареала. Оно происходит в результате того, что внутри одного ареала условия обитания могут различаться.
Так, среди растений примером может стать видовое разнообразие одуванчика, который различается на территории Евразии.
Относительный характер приспособленности кактуса
Растение демонстрирует удивительную способность выживания в суровейших условиях засухи: восковая пленка и колючки минимизируют испарение, хорошо развитая корневая система способна уходить глубоко в почву и накапливать влагу, иголки защищают от травоядных. Но, в ситуации проливных ливней, кактус погибает от переизбытка влаги из-за гниения корневой системы.
Относительный характер приспособленности белого медведя
На латыни этот медведь называется Ursus maritima, что означает морской медведь. Его шерсть отлично приспособлена к холодной воде.
Она не пропускает воду во время плавания и практически полностью задерживает отдачу тепла от кожного покрова животного.
Относительный характер приспособленности крота
Этот зверь обитает в основном в грунте. Имеет обтекаемую форму тела, мощные конечности лопатообразной формы с развитыми когтями. Он очень ловко роет многометровые тоннели.
И при этом совершенно не ориентируется на поверхности: его зрительная система неразвита, а перемещаться он может только ползком.
Относительный характер приспособленности верблюда
Горб верблюда – его гордость! Там скапливается драгоценная в условиях засухи вода. Конечно, не в прямом смысле вода, — это молекулы h3O, связанные с липидными, жировыми клетками.
Животное может долго переносить голод, лежать на раскаленном песке, у него минимизировано потоотделение. Не просто так кочевники Сахары передвигались именно на верблюдах. Но, увы, в заснеженных условиях этот выносливый красавец не справится с передвижением, питанием и поддержанием температуры тела.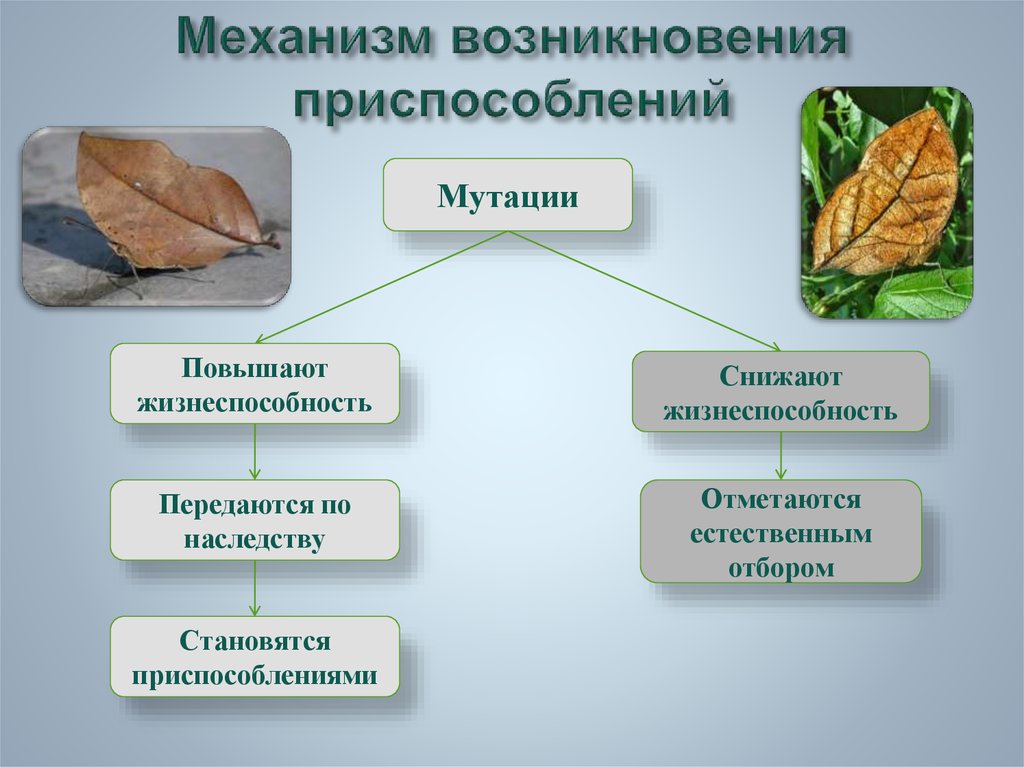
Чем характеризуется приспособленность растений к опылению насекомыми
Цветки растений прекрасны, непохожи друг на друга, ими хочется любоваться! Правда, биологическое значение этой красоты заключается вовсе не в том, чтобы радовать человека.
Главная задача цветущего растения – привлечь насекомого опылителя. Для этого используется несколько основных путей: яркая окраска крупных цветков, приятный для насекомых аромат, скученность мелких цветков в соцветия и, конечно, питательный нектар внутри цветка.
Вывод о приспособленности организмов к среде обитания
Выявление закономерностей и изучение приспособлений животного мира в различных формах наземной, водной, воздушной жизни является важной и безгранично интересной темой для исследователей. Поскольку раскрывает основные пути эволюционного процесса видоизменения живых существ.
Приспособленность организмов к условиям внешней среды и её относительный характер
Тема: Приспособленность организмов к условиям внешней среды и её относительный характер
Цель: изучение приспособленности организмов – при изучении темы получить
знания о механизмах возникновения и формирования приспособлений под
воздействием естественных факторов (наследственной изменчивости, борьбы за
существование, естественного отбора). Следует обосновать их совместное
действие, ведущую роль естественного отбора. На основе полученных знаний
необходимо выработать умение опровергать религиозные взгляды на
приспособленность.
Следует обосновать их совместное
действие, ведущую роль естественного отбора. На основе полученных знаний
необходимо выработать умение опровергать религиозные взгляды на
приспособленность.
Ход урока
Ко времени появления учения Дарвина об эволюции органического мира перед наукой встали три вопроса: (записать на доске)
1. Как объяснить приспособленность живых существ к среде?
2. Как возникло существующее в природе многообразие видов?
3. Почему в ходе эволюции происходит повышение организации живых существ?
Для их решения Дарвин обратился к практике сельского хозяйства. Он установил, что движущие силы пород домашних животных и сортов культурных растений – наследственная изменчивость и проводимый человеком отбор. Нам необходимо рассмотреть эти вопросы по отношению к видам, обитающим в естественных условиях.
I Объяснение приспособленности организмов в додарвиновский период
Крупный ученый конца 18 –
начала 19 века Кювье, и его ученики отстаивали взгляды о сотворении видов
Богом, о неизменяемости видов. Они предлагали теорию катастроф: на Земле
постоянно происходят катастрофы – они уничтожают всё живое. Ученики Кювье в
дальнейшем говорили о повторных актах творения: сначала Бог сотворил
примитивные организмы, потом катастрофа уничтожила, после чего Бог сотворил
более совершенные формы и современные виды.
Они предлагали теорию катастроф: на Земле
постоянно происходят катастрофы – они уничтожают всё живое. Ученики Кювье в
дальнейшем говорили о повторных актах творения: сначала Бог сотворил
примитивные организмы, потом катастрофа уничтожила, после чего Бог сотворил
более совершенные формы и современные виды.
Были высказаны представления о возможности превращения (трансформации) одного вида в другой при изменении условий среды.
Идеи о едином акте творения – креационизм – сторонник Карл Линней. По его мнению, каждый вид возник в результате акта творения Бога, а значит – виды неизменны и постоянны.
Провозвестником
эволюционизма был К. Линней, он придерживался идеи, что «видов столько, сколько
их создало Бесконечное существо». Историческая заслуга его состоит в том, что
он выдвинул принцип иерархичности систематических категорий (таксонов): виды
объединяются в роды, роды в семейства и пт. Иерархичность системы Линнея близка
к идее расхождения признаков у родственных организмов, предложенной столетием
назад Чарльзом Дарвином.
Сент. – Илер (1772 – 1844) – трансформист и эволюционист. Развил идею единого происхождения, идею единства плана строения. Он считал, что при изменении условий изменяются строение и функции организмов и весь организм в целом. Попытался доказать это экспериментально. Воздействуя различными факторами на яйцо птицы, и получив птичьих уродов, он как трансформист признал, что виды изменяются резко, что один вид при изменении условий может превратиться в другой.
Французский естествоиспытатель Ж. Б. Ламарк считал, что существующие виды должны со временем превратиться в новые, более совершенные. Но появление организации по Ламарку рассматривалось с идеалистических позиций, т. е повышение организации и организмов происходит при наличии и под воздействием некой нематериальной силы во всех организмах, которая и вызывает повышение уровня организации.
Ламарк выдвигал 2 причины эволюции:
1. Градация (повышение организации при нематериальных факторов)
2. Приспособительная дифференцировка (изменение органов под воздействием внешних условий среды)
Приспособленность к условиям внешней среды по Ламарку
Он
прибегает к теории флюидов – это тонкая невидимая материя, проводящая теплоту. В каждом местообитании свои условия, а значить имеются свои флюиды. Они
проникают в организм и заставляют его изменяться в соответствии с условиями
среды. Пример: У водного растения стрелолисты листья различаются – подводные
тонкие и рассеченные, надводные – лопастные. Это объясняется тем, что в Н2О
действуют одни флюиды, на воздухе — другие, в подводной среде – также отличные
от предыдущих.
В каждом местообитании свои условия, а значить имеются свои флюиды. Они
проникают в организм и заставляют его изменяться в соответствии с условиями
среды. Пример: У водного растения стрелолисты листья различаются – подводные
тонкие и рассеченные, надводные – лопастные. Это объясняется тем, что в Н2О
действуют одни флюиды, на воздухе — другие, в подводной среде – также отличные
от предыдущих.
Если изменяются условия внешней среды, у животных с нервной системой появляются новые потребности. Пример: климат стал более сухой – у животных появилась потребность часто пить.
С появлением новых потребностей возникают новые действия и привычки. При этом одни органы начинают усиленно упражняться, другие бездействуют, а некоторые и вовсе исчезают.
Ламарк формулирует два закона:
1. Органы, которые упражняются – усиливаются, те, которые не упражняются – ослабевают.
2. Все изменения, которые возникают под влиянием неупражнений или
упражнений – передаются по наследству. Для доказательства он приводит ряд
примеров:
Для доказательства он приводит ряд
примеров:
3. Кроты, слепыми живут под землей, орган зрения у них не упражняются, глаза плохо развиты, а у слепыша – отсутствуют.
4. У змей предки ползали на брюхе, не используя ног – это привело к редукции ног. Но змеи стремились пролезть через узкие отверстия – это привело к тому, что их тело стало узким и длинным.
5. У жирафов ноги были такими же, как у других травоядных, они попали в условия, где питание заключалось только в листьях. Именно это и привело их к увеличению длины шеи и ног.
По Ламарку в ходе эволюции могут возникать новые органы. Объяснил он это идеалистически, что внутренне чувство может вызвать возникновение новых органов. Пример: У самцов часто возникали приступы гнева, у них жидкость бросалась в голову, и следствием этого стали выросшие рога.
Вывод
Так как Ламарк считал, что любое изменение в
строении организма являлось полезным для организма, то оно способствовало приспособлению
в среде обитания. Эти идеи составляли наиболее слабую часть его учения. Заслуга
Ламарка состояла в том, что он определил, что организмы являются результатом
эволюции.
Эти идеи составляли наиболее слабую часть его учения. Заслуга
Ламарка состояла в том, что он определил, что организмы являются результатом
эволюции.
Итак, в додарвиновский период развития биологии приспособленность живых существ служила доказательством бытия божия: без всемогущего Творца сама природа не смогла так разумно устроить живые существа и так мудро приспособить их к среде
+
Налицо были противоречия между научными фактами, раскрывающими совершенство органического мира и предполагавшимися тогда объяснениями. Дело доходило до того, что, по словам К. А.Тимирязева обходились молчанием поразительные факты и даже просто отрицались, так как казались слишком изумительными, слишком раздражали ум, напоминая о его беспомощности объяснить их. Ключ к этой «незатейливой мучительной загадке» принес Дарвин.
5А II
Как
действовала эволюция вырабатывая те или иные приспособления. Как возникновение
этих приспособлений можно объяснить с точки зрения теории Дарвина.
Покажем это на примере формирования приспособлений у лопуха обыкновенного. Вы знаете, что это растение обитает около жилья человека. Допустим, что у предков лопуха не были развиты прицепки и росли они где нибудь в лугах. В силу каких то обстоятельств популяция оказалась около жилища человека. В результате непрерывного процесса возникновения мутаций, их комбинация при скрещивании, волны численности обеспечивали генетическую гетерогенность популяции. Поэтому растения отличались друг от друга рядом наследственных признаков:
1. размерами
2. отсутствием или наличием прицепок
Внутривидовая борьба за существование способствовала выживанию особей, у которых признак наличие прицепок был более развит, сформирован. В процессе естественного отбора именно эти особи оставляли плодовитое потомство и численность их в популяции возрастала.
Растения
нового поколения вновь несли разнообразные мутации. Среди мутаций могли быть и
такие, которые усиливали проявление отобранного ранее признака. Обладатели
этого признака вновь имели больше шансов выжить и оставить потомство. И так из
поколения в поколение на основе усиления, накапливания полезного
наследственного изменения совершенствовалась черта присполобленности лопуха
обыкновенного к распространению семян животными и человеком
Обладатели
этого признака вновь имели больше шансов выжить и оставить потомство. И так из
поколения в поколение на основе усиления, накапливания полезного
наследственного изменения совершенствовалась черта присполобленности лопуха
обыкновенного к распространению семян животными и человеком
Объяснение возникновения приспособленности, данное Ч. Дарвиным,в корне отличается от понимания этого процесса Ламарком, который выдвинул идею о врожденной способности организмов изменяться под влиянием среды только в полезную для них сторону.
Рассматривая другие примеры адаптаций в живой природе (Демонстрация гербариев) мы можем предположить, что механизм их возникновения общий: во всех случаях приспособления возникают не сразу в готовом виде как нечто данное, а длительно формируются в процессе эволюции посредством отбора особей, имеющих признак в наиболее выраженной форме
Делаем вывод в лабораторной работе и записываем вывод
Велик
мир живых существ, населяющих нашу планету. это и микроскопические организмы, и
животные огромных размеров, хищные или совсем безобидные. всего их на Земле 1,5
млн видов, между ними существует тесная взаимосвязь и сложное взаимодействие.
немало в природе животных, у которых нет ни быстрых ног, ни острых когтей или
крепких зубов, но которые хорошо приспособились к условиям жизни и имеют особые
средства защиты от врагов. Эти животные не могут обороняться агрессивным
способом, при опасности они стараются отпугнуть противника.
это и микроскопические организмы, и
животные огромных размеров, хищные или совсем безобидные. всего их на Земле 1,5
млн видов, между ними существует тесная взаимосвязь и сложное взаимодействие.
немало в природе животных, у которых нет ни быстрых ног, ни острых когтей или
крепких зубов, но которые хорошо приспособились к условиям жизни и имеют особые
средства защиты от врагов. Эти животные не могут обороняться агрессивным
способом, при опасности они стараются отпугнуть противника.
1) Одни выделяют неприятную пахучую или едкую жидкость;
2) Другие принимают неожиданную для врага позу устрашения;
3) Третьи отпугивают своей схожестью с несъедобными ядовитыми видами;
4) Для спасения жизни отдельные виды могут калечить себя — отбразывать кусок хвоста или ногу, за которые ухватится противник;
5) Есть животные, которые в момент опасности притворяются мертвыми;
6) Большое значение дл некоторых животных имеют покровительственная форма и окраска (защитная; сезонная; расчленяющая), благодаря которым они становятся малозаметными на окружающем фоне;
7) Ряд
видов обладает удивительными способностями: быстро изменять свою окраску,
маскироваться при помощи различных предметов, чувствовать приближение стихийных
бедствий и стараться избегать их.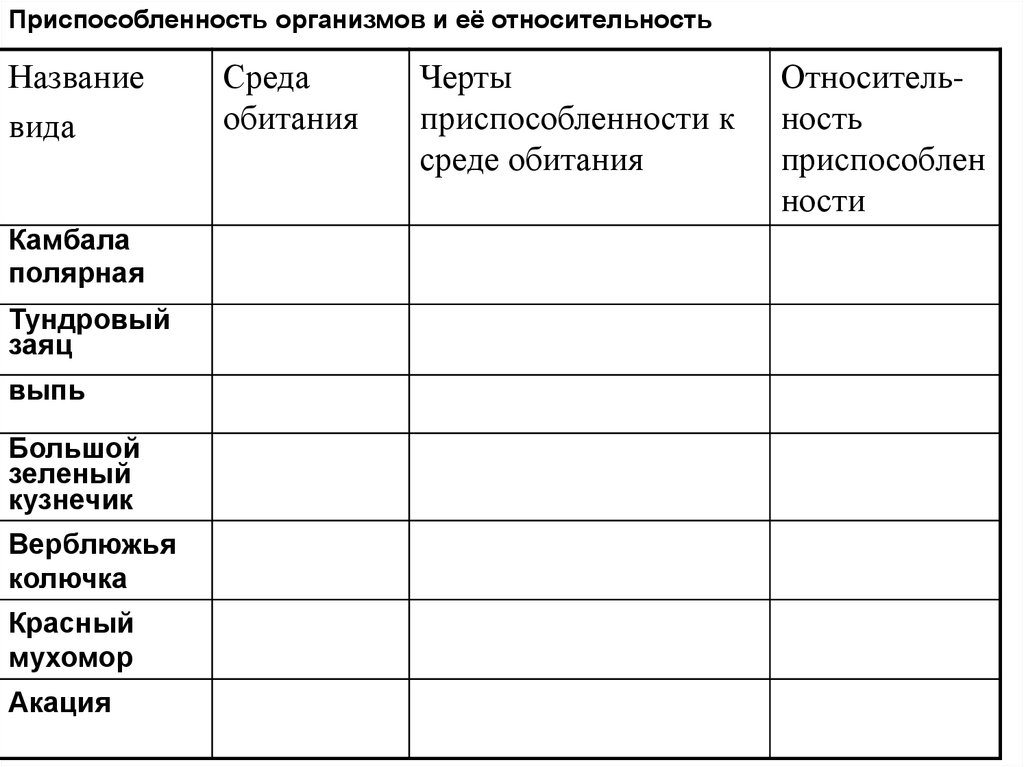
Приведите примеры приспособлений у животных
Примеры адаптаций
1. Защитная или покровительственная окраска.
Окраска животных имеет сходство с окраской среды, в которой они живут. Пример: пустынные змеи или ящереци окрашены в серо- желтый цвет под стать цвету окружающей почвы и растительности, а животные, обитающие среди мнегов, имеют белый мех или оперение.
2. У открытогнездящихся птиц глухарь, тетерев, сидящая на гнезде самка почти не отличима от фона, соответствует фону и пигментированная скорлупа . У пттиц, гнездящихся в дупле, самки имеют яркую окраску (прим. попугаи, дятлы)
Ночные бабочки днём могут погибнуть от насекомоядных птиц, но поскольку окраска их крыльев сливается с тоном той поверхности, на которой они проводят день, многие из них остаются незамеченными и переживают неблагоприятное для них время.
Окраска многих животных представляет собой сочетание
контрастных по цвету пятен и полос, которые не соответствуют форме животного,
но по тону и рисунку сливаются с окружающим фоном.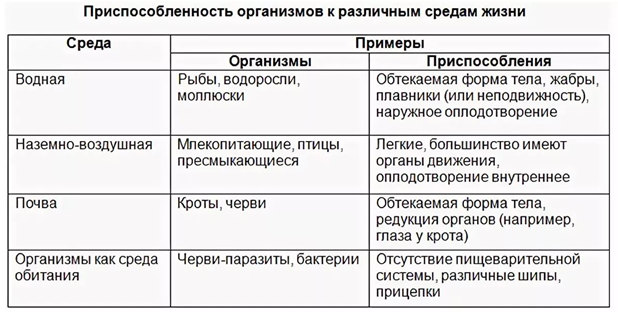 Расчленяющую окраску имеют
зебры и жирафы: их полосатые и пятнистые фигуры почти незаметны на фоне
растительности африканских саванн, особенно в сумерки, когда выходят на охоту
хищники. Такой вид окраски характерен и для обитателей подводного мира,
например рыбы ангела и вымпельных щетинозубов.
Расчленяющую окраску имеют
зебры и жирафы: их полосатые и пятнистые фигуры почти незаметны на фоне
растительности африканских саванн, особенно в сумерки, когда выходят на охоту
хищники. Такой вид окраски характерен и для обитателей подводного мира,
например рыбы ангела и вымпельных щетинозубов.
3. Отпугивающая окраска.
Животные с яркой окраской хорошо заметны на окружающем фоне, они не рпячутся — они несъедобны, ядовиты, их яркая окраска является предупреждением «Не трогай!» и характерна для жалящих насекомых, имеющих ядовитые железы, божьих коровок (птицы их не трогают)
4. Мимикрия.
Одним из защитных свойств животных является сходство беззащитного вида с хорошо защищенным. Среди насекомых широко распространено подражание жалоносным перепончатокрылым, есть бабочки-стеклянницы, напоминающие видом шершней.
Мимикрия — резутьтат отбора сходных мутаций у
различных видов. Она помогает незащищенным животным выжить, способствует
сохранению организма в борьбе за существование.
5. Меняющаяся окраска.
Природа наградила некоторых животных способностью изменять окраску при переходе из одной цветовой среды в другую. Такое свойство служит животному надежной защитой, поскольку делает его малозаметным в любой обстановке.
Меняют цвет древесные ящерицы — хамелеоны. быстрая смена окраски от зеленой к желтой или бурой делает её почти незаметной на ветках среди листвы. Кроме того, хамелеон может отпугивать врага быстрой сменой окраски контрастных цветов, становясь то ярко-зеленым, то красным иои черным.
6. Защитнаяф форма: Есть немало животных, тела которых похожи на какой либо предмет окружающей среды. такое сходство нередко спасает животное от врагов. Гусеницы некотрых бабочек напоминают сучки, а тело бабочек — лист или цветок.
7. Устрашающая поза.
Многие
животные, которые не обладают достаточной силой для отпора врагу, пытаются все же
отпугнуть его, принимая различные устрашающие позы. Например, ящерица ушастая
круглоголовка широко растягивает околоушные складки, которые наливаются кровью
и вместе со ртом создает впечатление огромной пасти.
8. Оборонительные выеделения.
Существует немало животных, которые при опасности «выстреливают» во врага какой либо жидкостью, обладающей защитными свойствами. Эти животные не имеют отпугивающей окраски, которая могла бы насторожить нападающего, и поэтому «выстрелы» неожиданными и эффективными. Осьминоги, кальмары — выпускают навстречу врагу «чернильную бомбу» — жидкость, нескольких капель которой достаточно чтобы замутить вокруг воду и незаметно скрыься.
9. Способность к прогнозированию.
Замечено,
что перед снихийным бедствием животные ведут себя беспокойно, но о
надвигающейся беде большинство из них «узнает» слишком поздно и не
успевает ее избежать. Но есть такие животные, которые «слышат»
приближение опасности с большим запасом времени и спасаются. Пример:
Приближение землетрясений змеи и крысы предчувствуют за 1,5 — 2 месяца; крысы
улавливают сигналы готовящегося землетрясения с эпицентра, находящегося от них
на расстоянии 100 и более километров.
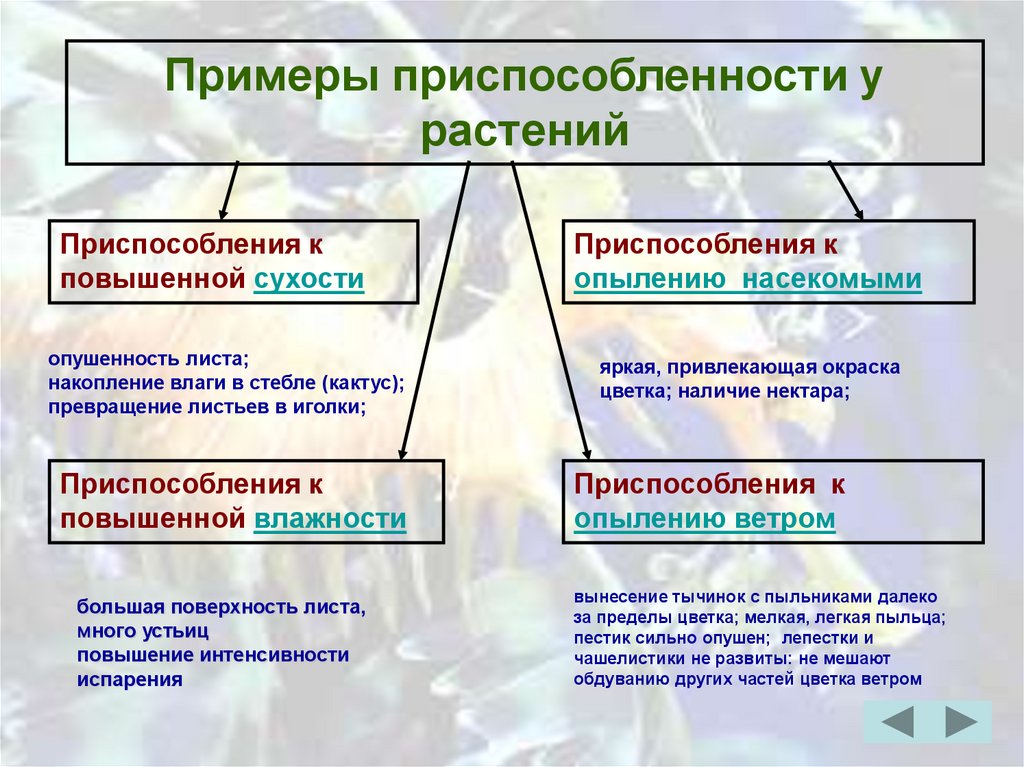
Какие вы знаете приспособления у растений?
Приспособленность у растений
Различные приспособления, помогающие растениям избегать вредного самоопыления, является одновременно и средством, обеспечивающим перекрестное опыление.
1. Образование различных цветков: пестичных или тычиночных. Пример: у огурца тычинковые цветы не приносят плодов. Их и называют поэтому пустоцветами.
2. Разновременное созревание тычинок и пестиков. Пример: у резеды пестик уже вытянется и раскроет рыльце для принятия пыльцы, а тычинки еще не этом растении еще не созрели и не могут опылить пестик. (иногда бывает наоборт)
Посмотрите, какое в природе разнообразие в формах и устройствах цветков. строение отдельных цветков приспособлено к строению и повадкам отдельных видов насекомых
3. Окраска цветков
приспособлена к особенностям зрения отдельных насекомых. Этой же цели служит и
строение цветочных венчиков. Пример: цветки полевой гвоздики представляют собой
удобную посадочную площадку для бабочек, а нектар скрыт в глубине венчика,
откуда его может достать только длинный хоботок тех же бабочек.
Этой же цели служит и
строение цветочных венчиков. Пример: цветки полевой гвоздики представляют собой
удобную посадочную площадку для бабочек, а нектар скрыт в глубине венчика,
откуда его может достать только длинный хоботок тех же бабочек.
4. Запах — тоже приспособление, возникшее в ходе эволюции.
5. Если приспособления широко рспространены, следовательно они являются полезными, т.к естественный отбор сохраняет только полезные.
6. Дарвин ставит эксперимент в двух вариантах для доказательства полезности перекрестного опыления:
· 1 вариант: растения посещались насекомыми, которые осуществляли перекрестное оплодотворение;
· 2 вариант: растения были покрыты тканью и насекомые не могли посещать их и осуществить перекрестное оплодотворение — там шло самоопыление.
Все условия были одинаковыми (температура, почва, вода). Результат: растения, которые не посещались насекомыми дали урожай семян намного меньше.
Еще большие различия
оказались в следующих поколениях. Растения, развивающиеся из семян
самоопыляющихся растений давали меньше зеленой массы, меньше цветов, семян,
растения эти были подвержены заболеваниям, гибли, особенно угнетающе это
сказалось в следующих поколениях. Дарвин вывел закон полезности
перекрестного оплодотворения.
Растения, развивающиеся из семян
самоопыляющихся растений давали меньше зеленой массы, меньше цветов, семян,
растения эти были подвержены заболеваниям, гибли, особенно угнетающе это
сказалось в следующих поколениях. Дарвин вывел закон полезности
перекрестного оплодотворения.
Половой отбор
Дарвин обратил внимание, что некоторые факты противоречат положениям об естественном отборе, т.е некоторые виды имеют яркую окраску, хорошо заметны, выделяются пением. Анализируя факты, Чарльз Дарвин создал представление о половом отборе. Это частный случай естественного отбора, происходящий в период размножения, свойственен особям одного пола. Пример: происходит между самцами за оплодотворение самки.
Биологическое значение
отбора состоит в стимуляции процессов размножения. в этом случае идет борьба
между самцами в форме боев на смерть. Самки в это время пассивны. Они достаются
победителю. эта форма отбора ведет к тому, что потомство появляется более
сильным, это ведет к увеличению у самцов органов, которые используются в
драках.
эта форма отбора ведет к тому, что потомство появляется более
сильным, это ведет к увеличению у самцов органов, которые используются в
драках.
Форма отбора идет в виде соперничества. Самка активно выбирает понравившегося самца. Самцы соперничают внешним видом, особенностями поведения, танцами, пением. Эта форма отбора к развитию вторичных половых признаков.
Относительный характер приспособленности.
Господствовало мнение, что каждое отдельное приспособление абсолютно, т.к соответствует определенной цели, заложенной творцом.
Приспособленность организмов к среде выработана в процессе длительного исторического развития под действием естественных причин, из большого разнообразия факторов, а значит и генотипов, в популяциях отбираются наиболее соответствующие данным условиям.
Лишь в результате очень
длительного эволюционногог процесса появляются организмы, хорошо адаптированные
к изменившимся условиям.
Поразительное сходство в строении органов, возникающее в ходе эволюции у очень отдаленных друг от друга форм животных и растений, свидетельствует в пользу сходным образом действовавшего естественного отбора. (Демонстрация гербариев)
Кроме естественного отбора и наследственной изменчивости в поддержании и формировании приспособлений существенную роль играют изолирующие механизмы, препятствующие широкому проникновению генетического материала из других видов.
Следует помнить, что приспособленность организмов к среде обитания не абсолютна, а относительна, так как условия среды обитания чато изменяются быстрее, чем формируются приспособления. Соответствуя конкретной среде обитания, приспособления теряют свое значение при ее изменении.
Доказательством относительного характера присобленности могут быть следующие факты:
1. Защитные приспособления от одних врагов оказываются неэффективными от других. Пример: ядовитых змей, опасных для многих животных, поедают мангусты, ежи)
2. Проявление инстинктов у животных может оказаться нецелесообразным
(ночные бабочки собирабт нектар со светлых цветков, хорошо заметных ночью, но
также летят и на огонь, хотя и гибнут при этом .
Проявление инстинктов у животных может оказаться нецелесообразным
(ночные бабочки собирабт нектар со светлых цветков, хорошо заметных ночью, но
также летят и на огонь, хотя и гибнут при этом .
3. Бобры, выбвигающие постройки в комнатках. Полезные в одних условиях органы становятся бесполезными и даже относительно вредными в другой среде (прим.: перепонки между пальцами у горных гусей, которые никогда не опускаются на воду).
4. Возможны и более совершенные приспособления к данной среде обитания. Некоторые виды животных и растений быстро размножались и широко распространяются в совершенно новых для них районах земного шара, куда были случайно или намеренно завезены.
5. Приспособленность организмов только к опредленной среде (рыбы, птицы).
6. Гибель организмов в результате узкой специализации (жало пчелы).
7. Ненадежность любой защиты ( яд среднеазиатского каракута безвреден для овец; иволга поедает мохнатых гусениц; лиса катит ежа в водоем, в котором его мускулатура расслабляется и иглы складываются)
8. Переход симбиоза в паразитизм ( взаимоотношения гриба и водоросли в
лишайнике).
Переход симбиоза в паразитизм ( взаимоотношения гриба и водоросли в
лишайнике).
Учение Дарвина об относительной приспособленности как результате естественногог отбора полностью опровергает существовавшие до него взгляды и утверждения о божественном происхождении организмов и абсолютном характере органической целесообразности (К. Линней), а также о врожденной способности организмов изменяться под влиянием среды только в полезную для них сторону (Ж.Б Ламарк)
Обобщение
· Приспособленность организмов к внешней среде и их органов к выполняемой ими функции является необходимым следствием естественного отбора;
· Приспособленность носит относительный характер
· Экспериментальнон доказательство естественного отбора.
Рис 7 стр 95 Методика
Рис 8 стр 96 увеличить
Сообщение стр 36 -37
Д/з
Стр 20
Задание для самостоятельной работы по биологии
Дидактический материал лекций посмтореть
Рисунки:
1. Стрелолист
Стрелолист
2. Крот, слепыш
3. Змея
4. Жираф
5. Олень с рогами
6.
Посмотреть гербарии, где есть приспособления у семян к распространению: ветром, саморазбрызгиванием, водой, человеком, птицами, животными
В чем разница между абсолютной и относительной приспособленностью?
Мне кажется, определение относительной и абсолютной пригодности не столь однозначно.
Абсолютная пригодность[править]
Абсолютная приспособленность (W) генотипа определяется как пропорциональная изменение численности этого генотипа в течение одного поколения обусловленное отбором. Например, если ${\displaystyle n(t)}$ обилие генотипа в поколении t в бесконечно большом популяции (чтобы не было генетического дрейфа), и пренебрегая изменение распространенности генотипов из-за мутаций, то[4] $ {\ Displaystyle п (т + 1) = Wn (т)} $. Абсолютная приспособленность больше 1 указывает на рост численности этого генотипа; абсолютная пригодность меньше 1 указывает на снижение.
Относительная пригодность[править]
Принимая во внимание, что абсолютная приспособленность определяет изменения в распространенности генотипов, относительная приспособленность (w) определяет изменения частоты генотипов. Если N(t) – общая численность населения в поколении t, а соответствующий частота генотипа равна $ {\ displaystyle p (t) = n (t) / N (t)} $, тогда $ {\ displaystyle p (t + 1) = {\ frac {w} {\ overline {w}}} p (t)} $, где ${\displaystyle {\overline {w}}}$ — средняя относительная приспособленность популяции (опять же без учета изменений частоты из-за дрейфа и мутация). Относительная приспособленность указывает только на изменение распространенности разных генотипов по отношению друг к другу, и поэтому только их значения относительно друг друга важны; относительная приспособленность может быть любое неотрицательное число, включая 0. Часто бывает удобно выбрать один генотип в качестве эталона и установить его относительную пригодность на 1.
Относительная пригодность используется в стандарте Райта-Фишера и Морана. модели популяционной генетики.
Абсолютные приспособленности можно использовать для расчета относительной пригодности, поскольку $ {\ displaystyle p (t + 1) = n (t + 1) / N (t + 1) = (W / {\ overline {W}}) p (t)} $ (мы использовали тот факт, что $ {\ displaystyle N (t + 1) = {\ overline {W}} N (t)} $, где ${\ displaystyle {\ overline {W}}} $ — среднее значение абсолютной пригодности. среди населения). Это означает, что $ {\ displaystyle w / {\ overline {w}} = W / {\ overline {W}}} $, или, другими словами, относительная приспособленность равна пропорциональна $ {\ displaystyle W / {\ overline {W}}} $. Это не можно вычислить абсолютную пригодность из относительной пригодности в одиночку, поскольку относительная пригодность не содержит информации об изменениях в общей численности населения

 Относительная пригодность используется в стандарте Райта-Фишера и Морана.
модели популяционной генетики.
Относительная пригодность используется в стандарте Райта-Фишера и Морана.
модели популяционной генетики.- Это хорошо, когда мы говорим о генотипах или фенотипах. Но
когда мы используем модели, например. фитнес-ландшафты, как мы можем
перевести это? Например, если у меня есть линейный фитнес-ландшафт
(сделано с линейной функцией), то значение «пригодности», которое я нахожу
с моделью «ожидаемая пригодность» или $\widehat y$. Таким образом, в этом
случае, это «пригодность всех возможных фенотипов, на основе на
популяционная приспособленность и фенотипическая информация». Называем ли мы это
относительное или абсолютное?
- Когда пригодность или один из ее компонентов вычисляется из модели (скажем, модель метки-повторного захвата), какой тип приспособленности вычисляется? В Марк Кери и Майкл Шауб (2012) Байесовский анализ населения с использованием WinBUGS: иерархическая перспектива , они показывают модели для вычисления вероятности выживания на основе истории повторной поимки. Это будет относительно или нет?
- В чем различие двух определений и не являются ли они слишком ограничительными, если мы хотим расширить их до моделей, вычисляющих приспособленность всех особей в популяции?
 фитнес-ландшафты, как мы можем
перевести это? Например, если у меня есть линейный фитнес-ландшафт
(сделано с линейной функцией), то значение «пригодности», которое я нахожу
с моделью «ожидаемая пригодность» или $\widehat y$. Таким образом, в этом
случае, это «пригодность всех возможных фенотипов, на основе на
популяционная приспособленность и фенотипическая информация». Называем ли мы это
относительное или абсолютное?
фитнес-ландшафты, как мы можем
перевести это? Например, если у меня есть линейный фитнес-ландшафт
(сделано с линейной функцией), то значение «пригодности», которое я нахожу
с моделью «ожидаемая пригодность» или $\widehat y$. Таким образом, в этом
случае, это «пригодность всех возможных фенотипов, на основе на
популяционная приспособленность и фенотипическая информация». Называем ли мы это
относительное или абсолютное? См. также Endler, J.A. (1986) Естественный отбор в дикой природе.
На странице 168 книги:
также Endler, J.A. (1986) Естественный отбор в дикой природе.
На странице 168 книги:
- Среднее абсолютное соответствие: $\overline {W} = [∑ƒ(X)W(X)]/[∑ƒ(X)]$
- ƒ(X) частота генотипа или фенотипа X
- Абсолютная пригодность: $W(X)$
- Относительная приспособленность: $w(X)= W(X) /\overline {W}$
- Таким образом, $\overline {w}(X)=1$
Относительную приспособленность также можно измерить по отношению к конкретному фенотипу (или генотипу), в котором cas $\overline {w}$ не обязательно равен 1; это наиболее распространенный метод, используемый для полиморфных признаков. Если популяция отбирается дважды (или более) в течение поколения, так что особи во второй выборке представляют собой подмножество особей, отобранных в первой выборке (как в исследовании «отлов-повторный отлов» или когортном исследовании), то можно рассчитать абсолютную пригодность. Примерами являются вероятность выживания между образцами или вероятность спаривания.
 С другой стороны, если образцы сделаны без замены или если образцы сделаны из молодых и взрослых особей одновременно, то можно рассчитать только относительную приспособленность; информация об общем количестве и средней пригодности теряется (см. обсуждение в O’Donald 1971, Хорнс и Харрисон, 1970, и Мэнли, 1974).)
С другой стороны, если образцы сделаны без замены или если образцы сделаны из молодых и взрослых особей одновременно, то можно рассчитать только относительную приспособленность; информация об общем количестве и средней пригодности теряется (см. обсуждение в O’Donald 1971, Хорнс и Харрисон, 1970, и Мэнли, 1974).)На странице 42 той же книги:
Приспособленность и приспособление. Адаптированность – это степень, в которой организм способен жить и размножаться в заданном наборе сред: состояние приспособленности (Добжанский, 1968а,б). Адаптация это процесс адаптации или более адаптации (там же). К сожалению, адаптация также используется в смысле адаптивного признака (Левонтин 1978), смешивая конечный продукт с процессом (см. также Данбар 1982). Адаптивная черта — это «аспект модели развития». организма, выживающего и размножающегося» (Добжанский, 1956, 1968а). Существуют проблемы с точным определением того, что такое адаптированный, чтобы его можно измерить (Добжанский 1956, 1968а,б; Стерн 1970; Левонтин 1978 год; Данбар 1982).
Бонус: я пытаюсь воссоздать уравнение из Ланде и Арнольда 1983. Но оно не работает. Я не знаю, что не так (строка, где вы видите (mean.rel.fit = mean(w.relative.fit)), этот результат должен дать 1, но это не в моем случае. Я не знаю почему):
# W: Абсолютная пригодность # w: относительная пригодность сет.
 2)
}
# Это не работает! Это считается нормальным
история (p.z (z),
перерывы = 30)
rnorm(nb.data,10,2)
# Линейно распределенная абсолютная фитнес-функция
Wz = функция (z) {
наклон = 1
перехват = 0
e=(z*наклон+пересечение)
(е-мин (г))/(макс (г)-мин (г))
}
# Существует линейная зависимость между чертой z и абсолютной приспособленностью W
график (z, W.z (z))
de = плотность (z,
п = 600000)
требуют (зоопарк)
# Если n = 600000 в функции плотности, то должно быть ~=1
сумма (diff (de $ x [порядок (de $ x)]) * rollmean (de $ y [порядок (de $ x)], 2))
дублируется (раунд (z, 1))
df.z = as.data.frame (таблица (округление (z, 1)))
значения = as.numeric(as.character(df.z$Var1))
frq = as.numeric(as.character(df.z$Freq))
frqcy = функция (длина) {frq [1: длина]}
f = функция (значения) (W.z (значения) * frqcy (длина (значения)))
(среднее.абсолютное.приспособленность = интегрировать(f,
подразделения = 2000,
отн.
2)
}
# Это не работает! Это считается нормальным
история (p.z (z),
перерывы = 30)
rnorm(nb.data,10,2)
# Линейно распределенная абсолютная фитнес-функция
Wz = функция (z) {
наклон = 1
перехват = 0
e=(z*наклон+пересечение)
(е-мин (г))/(макс (г)-мин (г))
}
# Существует линейная зависимость между чертой z и абсолютной приспособленностью W
график (z, W.z (z))
de = плотность (z,
п = 600000)
требуют (зоопарк)
# Если n = 600000 в функции плотности, то должно быть ~=1
сумма (diff (de $ x [порядок (de $ x)]) * rollmean (de $ y [порядок (de $ x)], 2))
дублируется (раунд (z, 1))
df.z = as.data.frame (таблица (округление (z, 1)))
значения = as.numeric(as.character(df.z$Var1))
frq = as.numeric(as.character(df.z$Freq))
frqcy = функция (длина) {frq [1: длина]}
f = функция (значения) (W.z (значения) * frqcy (длина (значения)))
(среднее.абсолютное.приспособленность = интегрировать(f,
подразделения = 2000,
отн. .05,
нижний = мин (г),
верхний = макс (г))
)
# Относительная пригодность
w.z <- функция (z) {
W.z(z)/mean.absolute.fitness$значение
}
(w.relative.fit = w.z(z))
# Это должно быть 1
(среднее значение относительного соответствия = среднее значение (w.относительное соответствие))
w.relative.fit
f2 = функция (z) (z * w.z (z) * p.z (z))
(среднее.абсолютное.приспособленность = интегрировать(f2,
нижний = мин (г),
верхний = макс (г))
)
.05,
нижний = мин (г),
верхний = макс (г))
)
# Относительная пригодность
w.z <- функция (z) {
W.z(z)/mean.absolute.fitness$значение
}
(w.relative.fit = w.z(z))
# Это должно быть 1
(среднее значение относительного соответствия = среднее значение (w.относительное соответствие))
w.relative.fit
f2 = функция (z) (z * w.z (z) * p.z (z))
(среднее.абсолютное.приспособленность = интегрировать(f2,
нижний = мин (г),
верхний = макс (г))
)
Путают ли когда-нибудь экологи абсолютную и относительную приспособленность?
Начну с короткой версии этого поста, в виде старого анекдота. Потом уточню.
Вот короткая версия: Двое туристов идут по лесу. Они поворачивают и видят медведя на тропе впереди них. Медведь бросается на них. Они начинают бежать, но медведь их догоняет. Внезапно первый турист садится и начинает менять походные ботинки на кроссовки. Второй турист говорит: «Что ты делаешь?! Ты никогда не убежишь от этого медведя, в кроссовках или без кроссовок!» Первый турист отвечает: «Мне не нужно убегать от медведя, мне нужно убежать от тебя». 🙂
Второй турист говорит: «Что ты делаешь?! Ты никогда не убежишь от этого медведя, в кроссовках или без кроссовок!» Первый турист отвечает: «Мне не нужно убегать от медведя, мне нужно убежать от тебя». 🙂
Шутка о разнице между абсолютной и относительной приспособленностью и о важности их не смешивать. В эволюции обычно интерес представляет относительная приспособленность, поскольку это все, о чем заботится естественный отбор. Например, даже если два аллеля в одном и том же локусе оба не подходят в каком-то абсолютном смысле, частота более подходящего из двух все равно будет увеличиваться за счет другого. То же самое верно и в области экологии сообщества. Если вы пытаетесь понять такие вопросы, как конкурентное исключение, сосуществование и сохранение разнообразия, то важны относительные способности разных видов к выживанию и размножению. Например, даже если вид выживает и хорошо размножается в каком-то абсолютном смысле, он будет исключен, если конкурирующий вид выживает и размножается еще лучше.
Экологи когда-нибудь путают абсолютную и относительную приспособленность? Я не уверен. Я никогда не сталкивался с действительно явным случаем смешения двух вещей, но иногда сталкивался со случаями, которые кажутся двусмысленными. Надеюсь, читатели присоединятся к другим примерам или прояснят мои недоразумения в примерах, которые я обсуждаю ниже (все примеры взяты из областей литературы, с которыми я не очень хорошо знаком).
Самая близкая к очевидной путанице, о которой я знаю, исходит из специализированной литературы по строительству. В рецензии на книгу философ биологии Самир Окаша уловил ведущих сторонников построения ниши, Лаланда и др., в несоответствии между их словесным определением «позитивного» построения ниши и их математическими моделями этого явления. Их словесное определение было основано на относительной пригодности, но их математические модели были основаны на абсолютной пригодности. Лаланд и др. признал двусмысленность. Но не читая книгу, о которой идет речь, я не могу комментировать дальше.
Окаша сравнивает оговорку Лаланда и др. с неспособностью отличить «сильный» альтруизм от «слабого» в эволюции. Сильный альтруизм относится к действиям, которые снижают абсолютную приспособленность организма и повышают абсолютную приспособленность других. Слабый альтруизм относится к действиям, которые повышают абсолютную приспособленность организма, одновременно повышая абсолютную приспособленность других организмов, тем самым снижая относительную приспособленность альтруистов по сравнению с неальтруистами. Слабый альтруизм не может развиться в неструктурированной панмиктической популяции. Но я иногда видел, как экологи заявляли, что экологический эквивалент может появиться. Я говорю «появляется», потому что словесные аргументы, которые я видел, кажутся мне двусмысленными, и поэтому я не совсем понимаю, что именно утверждается.
Например, растения, растущие в определенных «суровых» условиях, таких как альпийские луга, обычно способствуют, а не конкурируют со своими соседями, а это означает, что удаление соседей фокального растения снижает, а не увеличивает выживаемость, рост и/или плодовитость взрослых особей. То есть соседи увеличивают (некоторые компоненты) абсолютную приспособленность друг друга*. Это побудило некоторых авторов размышлять о том, делает ли фасилитация между растениями «реализованную нишу» больше, чем «фундаментальную нишу», и позволяет ли некоторым видам сохраняться в окружающей среде. где они иначе не могли. Как я уже сказал, это предположение неоднозначно; не все возможные варианты могут работать, я не думаю.** В частности, только потому, что ваши соседи поднимают ваши абсолютная пригодность означает, что , а не , они не могут исключить вас из соревнований. Их относительная пригодность все еще может быть выше вашей — возможно, отчасти потому, что вы также помогаете им!
То есть соседи увеличивают (некоторые компоненты) абсолютную приспособленность друг друга*. Это побудило некоторых авторов размышлять о том, делает ли фасилитация между растениями «реализованную нишу» больше, чем «фундаментальную нишу», и позволяет ли некоторым видам сохраняться в окружающей среде. где они иначе не могли. Как я уже сказал, это предположение неоднозначно; не все возможные варианты могут работать, я не думаю.** В частности, только потому, что ваши соседи поднимают ваши абсолютная пригодность означает, что , а не , они не могут исключить вас из соревнований. Их относительная пригодность все еще может быть выше вашей — возможно, отчасти потому, что вы также помогаете им!
Экономист Ник Роу провел аналогию, которая с небольшими изменениями прекрасно иллюстрирует этот момент. Представьте, что вы сидите в переполненном театре и не можете видеть сцену. Так что вы встаете, и тогда вы можете видеть лучше. Но это не значит, что всем будет лучше видно, если все встанут, или что вам лично будет лучше видно, если все встанут. Мы можем немного видоизменить эту аналогию, чтобы объяснить, почему межвидовое облегчение, наблюдаемое в альпийских растительных сообществах, не обязательно позволяет данному виду растений расти там, где иначе он не мог бы расти. Представьте себе людей, сидящих в переполненном театре. Под каждым сиденьем есть рычаг, который поднимает сиденье. Но никто не может дотянуться до рычага под своим сиденьем; добраться до него могут только их соседи. Если каждый дергает за чужой рычаг (т. е. каждый повышает абсолютную пригодность своих соседей), это не значит, что все в театре смогут видеть лучше. Действительно, может быть, никто не увидит лучше, или кто-то увидит лучше, а кто-то хуже. Поэтому, если вас интересует, как фасилитация влияет на сосуществование растений или их распределение по градиентам окружающей среды, вам нужно составить модель, определяющую, как фасилитация влияет на относительную, а не (только) абсолютную приспособленность разных видов (точнее, ваша модель должны объяснить, как фасилитация делает относительную приспособленность отрицательно частотно-зависимой).
Мы можем немного видоизменить эту аналогию, чтобы объяснить, почему межвидовое облегчение, наблюдаемое в альпийских растительных сообществах, не обязательно позволяет данному виду растений расти там, где иначе он не мог бы расти. Представьте себе людей, сидящих в переполненном театре. Под каждым сиденьем есть рычаг, который поднимает сиденье. Но никто не может дотянуться до рычага под своим сиденьем; добраться до него могут только их соседи. Если каждый дергает за чужой рычаг (т. е. каждый повышает абсолютную пригодность своих соседей), это не значит, что все в театре смогут видеть лучше. Действительно, может быть, никто не увидит лучше, или кто-то увидит лучше, а кто-то хуже. Поэтому, если вас интересует, как фасилитация влияет на сосуществование растений или их распределение по градиентам окружающей среды, вам нужно составить модель, определяющую, как фасилитация влияет на относительную, а не (только) абсолютную приспособленность разных видов (точнее, ваша модель должны объяснить, как фасилитация делает относительную приспособленность отрицательно частотно-зависимой). Что некоторые люди и начинают делать, хотя это только начало, и я не думаю, что очень широкий спектр моделей уже изучен.
Что некоторые люди и начинают делать, хотя это только начало, и я не думаю, что очень широкий спектр моделей уже изучен.
Последний контекст, в котором я задаюсь этим вопросом, касается сравнения того, насколько хорошо интродуцированные виды ведут себя в своих естественных и интродуцированных ареалах. У меня сложилось впечатление, что многие статьи рассматривают абсолютную приспособленность или ее компоненты интродуцированных видов в их естественных и интродуцированных ареалах. Например, спрашивая, имеет ли интродуцированный вид растений более высокую выживаемость, рост или плодовитость в своем интродуцированном ареале, чем в его естественном ареале. Никогда не понимал, почему такое сравнение интересов. Например, что, если интродуцированный ареал — просто лучшее место для произрастания всех растений, так что интродуцированный вид имеет более высокую абсолютную приспособленность в своем интродуцированном ареале, чем в своем естественном ареале, но более низкую относительную приспособленность?
Аналогия с эволюцией помогает прояснить мой вопрос. Биологи-эволюционисты стараются различать два различных значения термина «местная адаптация» (хороший обзор см. в Kawecki and Ebert 2004). Одно чувство, иногда называемое «дом против отсутствия», сравнивает абсолютную приспособленность данного генотипа на его родном («домашнем») участке с каким-либо другим («вне дома») местом, где он обычно не встречается. Но это ощущение локальной адаптации согласуется с тем, что отбор повсеместно отдает предпочтение одному и тому же генотипу. Если вы хотите понять пространственную изменчивость в отборе, вам необходимо изучить локальную адаптацию в смысле сравнения «домашнее против чужого». То есть нужно измерить приспособленность генотипов, растущих на их домашнем участке относительно к приспособленности чужеродных генотипов, которые были пересажены в это место. Если вы повторите это для нескольких участков и обнаружите, что на каждом участке местные генотипы всегда имеют более высокую относительную приспособленность , чем чужеродные, то вы продемонстрировали «местную адаптацию» в эволюционно значимом смысле.
Биологи-эволюционисты стараются различать два различных значения термина «местная адаптация» (хороший обзор см. в Kawecki and Ebert 2004). Одно чувство, иногда называемое «дом против отсутствия», сравнивает абсолютную приспособленность данного генотипа на его родном («домашнем») участке с каким-либо другим («вне дома») местом, где он обычно не встречается. Но это ощущение локальной адаптации согласуется с тем, что отбор повсеместно отдает предпочтение одному и тому же генотипу. Если вы хотите понять пространственную изменчивость в отборе, вам необходимо изучить локальную адаптацию в смысле сравнения «домашнее против чужого». То есть нужно измерить приспособленность генотипов, растущих на их домашнем участке относительно к приспособленности чужеродных генотипов, которые были пересажены в это место. Если вы повторите это для нескольких участков и обнаружите, что на каждом участке местные генотипы всегда имеют более высокую относительную приспособленность , чем чужеродные, то вы продемонстрировали «местную адаптацию» в эволюционно значимом смысле. То есть вы продемонстрировали пространственную изменчивость отбора, при этом разные генотипы отдавали предпочтение отбору в разных местах. Аналогичным образом, если вас интересует, почему численность интродуцированного вида быстро увеличивается в его интродуцированном ареале и вытесняет там местные виды, не следует ли вам спросить, выше ли его относительная приспособленность, когда он находится в «чужом» (т.е. интродуцированном) ареале? ) сайт, чем на своем «домашнем» (родном) сайте? Не спрашивая о его абсолютной пригодности «дома» и «в гостях»? Но я совсем не знаком с литературой по интродуцированным видам, так что, может быть, я просто совершенно не понимаю суть исследований, о которых говорю?
То есть вы продемонстрировали пространственную изменчивость отбора, при этом разные генотипы отдавали предпочтение отбору в разных местах. Аналогичным образом, если вас интересует, почему численность интродуцированного вида быстро увеличивается в его интродуцированном ареале и вытесняет там местные виды, не следует ли вам спросить, выше ли его относительная приспособленность, когда он находится в «чужом» (т.е. интродуцированном) ареале? ) сайт, чем на своем «домашнем» (родном) сайте? Не спрашивая о его абсолютной пригодности «дома» и «в гостях»? Но я совсем не знаком с литературой по интродуцированным видам, так что, может быть, я просто совершенно не понимаю суть исследований, о которых говорю?
*Это предостережение относительно фитнес-компонентов на самом деле очень важно. Тот факт, что альпийские растения обычно увеличивают выживаемость, рост и/или плодовитость взрослых особей своих соседей, означает ли , а не , что они на самом деле повышают, а не понижают абсолютную приспособленность своих соседей, поскольку семена должны прорасти, а сеянцы должны дожить до взрослой жизни. чтобы семенное растение завершило свой жизненный цикл. В самом деле, трудно представить себе систему, в которой каждый индивид действительно увеличивает абсолютную приспособленность своих соседей. В такой системе все абсолютные приспособленности будут положительно зависеть от плотности, а общая плотность резко возрастет до бесконечности. Но вполне возможно представить себе систему, в которой, скажем, соседи увеличивают выживаемость и плодовитость друг друга, но на самом деле снижают приспособленность друг друга, занимая участки, которые в противном случае могли бы быть заняты новыми потомками других особей. Моя цель здесь не в том, чтобы критиковать эмпирические исследования фасилитации на альпийских растениях и других системах только за измерение определенных компонентов физической подготовки. Я просто делаю здесь концептуальный вывод, который, я подозреваю, знаком многим читателям (включая тех, кто занимается фасилитации), но, возможно, не всем.
чтобы семенное растение завершило свой жизненный цикл. В самом деле, трудно представить себе систему, в которой каждый индивид действительно увеличивает абсолютную приспособленность своих соседей. В такой системе все абсолютные приспособленности будут положительно зависеть от плотности, а общая плотность резко возрастет до бесконечности. Но вполне возможно представить себе систему, в которой, скажем, соседи увеличивают выживаемость и плодовитость друг друга, но на самом деле снижают приспособленность друг друга, занимая участки, которые в противном случае могли бы быть заняты новыми потомками других особей. Моя цель здесь не в том, чтобы критиковать эмпирические исследования фасилитации на альпийских растениях и других системах только за измерение определенных компонентов физической подготовки. Я просто делаю здесь концептуальный вывод, который, я подозреваю, знаком многим читателям (включая тех, кто занимается фасилитации), но, возможно, не всем.
**Одним из способов (не обязательно единственным) это предположение может сработать, если у вас есть один вид (назовем его видом X), который помогает своим соседям, но не подвергается их влиянию.
Leave A Comment