
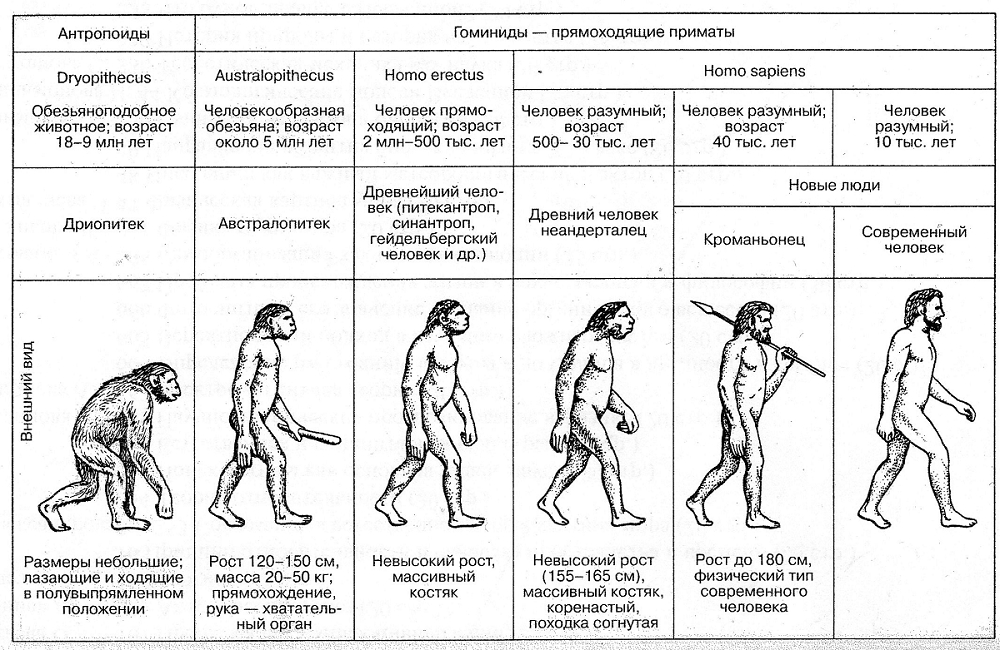
Тест по биологии на тему «Макроэволюция» (11 класс)
Закономерности и пути развития органического мира.
Макроэволюция и её доказательства
Вариант 1
1) Биологический регресс характеризуется
A. Увеличением численности особей данного вида
B. Возрастанием численности особей данного вида
C. Сужением ареала
D. Возрастанием числа видов, подвидов, популяций
2) Видообразование – это результат
A. Микроэволюции
B. Макроэволюции
C. Естественного отбора
D. Борьбы за существование
3) Победа вида или другой систематической группы в борьбе за существование
A. Прогресс
B. Дрейф генов
C. Ароморфоз
D. Регресс
4) Идиоадаптацией называют…
A. Приспособление организмов к условиям среды без перестройки уровня организации
B.
C. Резкое упрощение организации связанное с исчезновением целых систем органов
D. Все ответы верны
5) К ароморфозам следует отнести
A. Яркие цветки насекомоопыляемых растений
B. Возникновение защитной окраски
C. Появление фотосинтеза
D. Возникновение мимикрия
6) Ароморфоз иначе называется —
A. Аллогенез
B. Микроэволюция
C. Арогенез
D. Регресс
7) Конвергенция – это
A. Расхождение признаков в процессе эволюции
B. Сходные признаки у неродственных групп
C. Объединение нескольких популяции в одну
D. Образование изолированной группы внутри популяции
8) К идиоадаптации следует отнести
A. Появление полового процесса
B. Покровительственную окраску
C. Появление фотосинтеза
D. Лёгочное
дыхание
Лёгочное
дыхание
9) Примером
дегенерации можно считать отсутствие:
A.
Конечностей у змей
B. Зубов у
птиц
C. Стебля у
ряски
D. Жабр у
взрослых лягушек
10) Какое из перечисленных приспособлений не является ароморфозом?
A. Возникновение позвоночника у хордовых
B. Возникновение хобота у слона
C. Образование 2-х кругов кровообращения
D. Образование 3-х камерного сердца у земноводных
11) Авторы биогенетического закона
A. Фокс, Мюллер
B. Э.Геккель, Ф.Мюллер
C. Мюллер, Холдейн
D. Северцев, Шмальгаузен
12) Как называется расхождение в ходе эволюции признаков и свойств у первоначально близких групп организмов?
A. Дивергенция
B. Конвергенция
C. Адаптация
D. Изоляция
13) Направление эволюции, связанное с упрощением строения и образа жизни, называется
A. Идиоадаптацией
Идиоадаптацией
B. Дегенерацией
C. Дивергенцией
D. Ароморфозом
14) Семя, возникающее
у голосеменных растений, можно рассматривать как:
A.
Ароморфоз
B.
Идиоадаптацию
C.
Дегенерацию
D.
Дивергенцию
15) Аналогичными органами являются
A. Усики винограда и усики огурца
B. Крылья бабочки и крылья птицы
C. Иглы дикобраза и иглы ежа
D. Колючки кактуса и колючки барбариса
16) Одним из результатов макроэволюции является образование новых
A. Особей
B. Классов
C. Видов
D. Популяций
17) Кто установил филогенетические ряды?
A. А.Н.Северцов
B. В.О.Ковалевский
C. Ж.Б.Ламарк
D. К.Ф.Рулье
18) Макроэволюция:
A. Совокупность эволюционных процессов, происходящих внутри вида
B. Происходит
быстро и внезапно
Происходит
быстро и внезапно
C. Надвидовая эволюция
D. Доступна для наблюдения
19) Решающим эмбриологическим доказательством эволюции является
A. Сходство деления клеток у всех организмов
B. Сходство в строении скелетов млекопитающих разных отрядов
C. Сходство ранних стадий развития зародышей разных классов
D. Общность строения кровеносной системы млекопитающих
20) Признаки, каких классов сочетает в себе утконос?
A. Рептилий и млекопитающих
B. Птиц и рептилий
C. Птиц и млекопитающих
D. Амфибий и рептилий
21) Какие из перечисленных органов являются гомологичными?
A. Жабры рака и легкие кошки
B. Хобот слона и рука человека
C. Лапа крота и лапа обезьяны
D. Глаз кальмара и глаз млекопитающего
22) Биогенетический закон гласит:
A. Органический
мир развивается
Органический
мир развивается
B. Движущей силой эволюции является естественный отбор
C. Онтогенез кратко повторяет филогенез
D. Материальными носителями наследственной информации являются гены, локализованные в хромосомах
23) В результате ароморфоза образуются
A. Виды
B. Семейства
C. Классы
D. Роды
24) Сравнительная анатомия изучает
A. Ископаемые остатки растений и животных
B. Общность и различия в строении организмов
C. Сходство зародышей разных организмов
D. Общность и различия в функция организмов
25) «Живые ископаемые»
A. Утконос
B. Археоптерикс
C. Мамонт
D. Зверозубые ящеры
Закономерности и пути развития органического мира.
Макроэволюция и её доказательства
Вариант 2
1) Биологический прогресс характеризуется
A.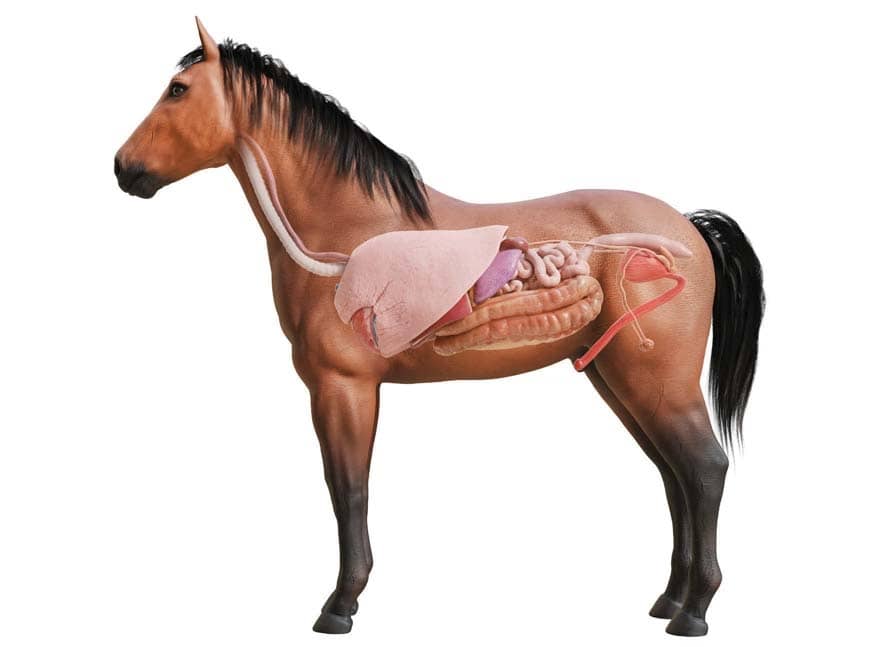
B. Возрастанием численности особей данного вида
C. Сужением ареала
D. Уменьшением числа видов, подвидов, популяций
2) В результате макроэволюции образуется
A. Вид
B. Подвид
C. Класс
D. Популяция
3) Эволюционное направление, которое характеризуется проявлением у организмов мелких приспособительных признаков —
A. Идиоадаптация
B. Дивергенция
C. Дегенерация
D. Ароморфоз
4) Гомологичными органами считают:
A. Сходные по происхождению
B. Выполняющие сходные функции
C. Не имеющие общего плана строения
D. Различные по происхождению
5) Дивергенция – это
A. Расхождение признаков в процессе эволюции
B. Схождение признаков
C. Объединение нескольких популяции в одну
D. Образование
изолированной группы внутри популяции
Образование
изолированной группы внутри популяции
6) Примером
конвергенции можно считать черты сходства между дельфином и:
A. Тюленем
B. Акулой
C. Синим китом
D. Всеми этими животными
7) Примером
идиоадаптации можно считать:
B. Два круга кровообращения у земноводных
C. Усики у гороха
D. Вторая сигнальная система у человека
8) Примером
ароморфоза можно считать:
A. Красивый
хвост у павлина
B. Крепкий
клюв у дятла
C. Длинные
ноги у аиста
D. Перья у
птицы
9) Отсутствие
кишечника у бычьего цепня, можно рассматривать как:
A.
Ароморфоз
B.
Идиоадаптацию
C.
Дегенерацию
D.
Конвергенцию
10) Какое из перечисленных приспособлений относят к идиоадаптациям?
A. Возникновение хорды
B. Возникновение ползучего стебля у клубники
C. Образование 2-х кругов кровообращения
D. Утрата
органов кровообращения у бычьего цепня
Утрата
органов кровообращения у бычьего цепня
11) Опыление цветков насекомыми и ветром — это пример …
A. Дегенерации
B. Идиоадаптации
C. Ароморфоза
D. Регресса
12) Появление у растений листьев, стеблей, корня это пример
A. Дегенерация
B. Идиоадаптация
C. Ароморфоз
D. Регресс
13) Отсутствие
выделительной системы у печёночного сосальщика можно рассматривать как:
A.
Ароморфоз
B.
Идиоадаптацию
C.
Дегенерацию
D.
Дивергенцию
14)
A. Разная форма клюва у галапагосских вьюрков
B. Белая окраска оперения у тундровой куропатки и шерсти у зайца-беляка
C. толстый слой подкожного жира и ласты у морского котика
D. различные способы опыления цветков у покрытосеменных
15) Гомологичные органы
A. Жабры рыбы и жабры рака
B. Колючки
кактуса и колючки боярышника
Колючки
кактуса и колючки боярышника
C. Усики гороха и усики винограда
D. Волосы млекопитающих и перья птиц
16) Какие доказательства эволюции основываются на изучении ископаемых остатков
A. Морфологические
B. Эмбриологические
C. Палеонтологические
D. Биогеографические
17) Какие организмы являются переходными формами?
A. Кистепёрые рыбы
B. Семенные папоротники
C. Археоптерикс
D. Прыткая ящерица
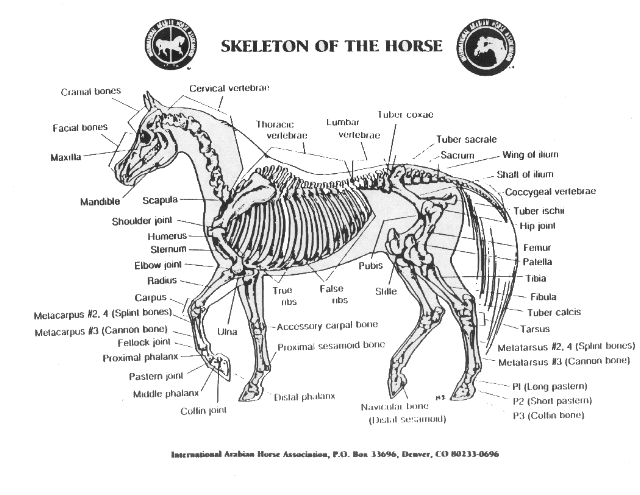
18) Какие органы лошадей претерпевали наибольшие изменения?
A. Конечности
B. Сердце
C. Пищеварительный тракт
D. Размеры тела
19) Кто сформулировал биогенетический закон?
A. Ч.Дарвин
B. А.Н.Северцов
C. Ф.Мюллер, Э.Геккель
D. К.Линней
20) Аналогичные органы:
A. Передняя конечность летучей мыши и кита
B. Передняя
конечность лягушки и крыло птицы
Передняя
конечность лягушки и крыло птицы
C. Крыло бабочки и крыло птицы
D. Усики гороха и колючки кактуса
21) К идиоадаптациям у голосеменных растений относят
A. Появление спор
B. Образование семени
C. Образование проводящих тканей
D. Видоизменение листьев
22) Учёный, изучивший историю развития лошади
A. А.Н.Северцов
B. В.О.Ковалевский
C. Ж.Б.Ламарк
D. К.Ф.Рулье
23) В результате ароморфоза образуются
Урок «Пройдём путём эволюции»
Ход
Эволюция – необратимый процесс исторического изменения живого (филогенез).
Попытки понять, как возникли живые организмы на Земле, почему они так разнообразны, появились у людей в далёкой древности. Долгое время господствовали религиозные взгляды на природу, которые утверждают, что все организмы созданы Богом и существуют в неизменяемом виде.
Основоположником научного объяснения – учения об историческом развитии органического мира- является английский натуралист XIX века Чарльз Дарвин.
Слайд 1 – 2. Сегодня мы отправляемся на экскурсию в Дарвинский Государственный музей в Москве, где организована выставка: «Пройди путём эволюции».
Учащиеся получают «билеты» — маршрутные листы, где они могут работать в ходе урока.
Слайд3. Жизнь и деятельность Ч. Дарвина – выступление учащегося. Основные вехи жизни и деятельности Дарвина:
12 февраля 1809 г, Шрусбери, родился Ч. Дарвин
Учёба в Эдинбургском университете на медицинском факультете, далее в Кембридже на богословском факультете.
1831-1836г.г. Кругосветное путешествие на корабле «Бигль», удивительные находки: мегатерий, зуб американской лошади, фауна Галапагосских островов…
1859год – вышел труд «Происхождение видов путём естественного отбора или сохранение благоприятствуемых пород в борьбе за жизнь
19 апреля 1882г Дарвина не стало.
Слайд 4-6. И этот гипс, заполнив пустоту,
Застыл и принял очертанья тела,
Которое давно уже истлело
Э. Дарвин
Удивительное сходство в строении и функционировании живых организмов не было следствием случайного совпадения. Оно является результатом их общего происхождения и указывается на единый «корень» жизни на нашей планете.
Оно является результатом их общего происхождения и указывается на единый «корень» жизни на нашей планете.
Отдел I Палеонтологические доказательства. Выступление учащегося – «экскурсовода». Демонстрация ископаемых останков. Основные понятия: палеонтология, ископаемые останки, окаменелости, ископаемые формы, эволюционные ряды.
К слайду 6 – см. дополнительную информацию.
Слайд 7-8. Кроме того, посмотри на людей,
Посмотри на немое
Племя чешуйчатых рыб, на стада,
На зверей и на стаи
Пёстрые птиц…
Лукреций Кар
Отдел II. Эмбриологические доказательства. Выступление «экскурсовода».
Факт единства происхождения живых организмов был установлен на основе эмбриологических исследований, в основе которых лежат данные науки эмбриологии.
Работа с материалом из книги .
Вопрос: Что пишет Дарвин по поводу этого вопроса?
Слайд 9. В борьбе стихий, в развитье постепенном
Все существа, все формы создались
И жизнею могучею зажглись!
Э. Дарвин
Дарвин
Отдел III.
Сравнительно-анатомические доказательства. Сравнительная анатомия – наука, изучающая и сравнивающая внутреннее и внешнее строение живых организмов.
Гомологичные органы- органы, имеющие общее происхождение, могут выполнять различные функции.
Аналогичные органы – органы, имеющие различное происхождение, но могут выполнять сходные функции
Работа с материалом из книги .
Вопрос: Что пишет Дарвин по поводу этого вопроса?
Слайд 10. Знакомство ещё с одной группой сравнительно-анатомических доказательств.
Рудименты – недоразвитый, остаточный орган, бывший полноценным на предшествующих стадиях развития.
Атавизмы – появление у потомков признаков, свойственных их отдалённым предкам.
Работа с материалом из книги .
Вопрос: Что пишет Дарвин по поводу этого вопроса?
Подведение итогов «экскурсии»
Выполнить тестовое задание в «билетах».
Домашнее задание: §49. Проверить решение теста.
Задания на выбор.
1. Используя материалы Интернет и других источников найти филогенетические ряды животных.
2. Подумайте, какие полезные ископаемые нашей местности могли бы быть достойными экспонатами музея, в каком отделе? (Можно собрать коллекцию )
Приложения
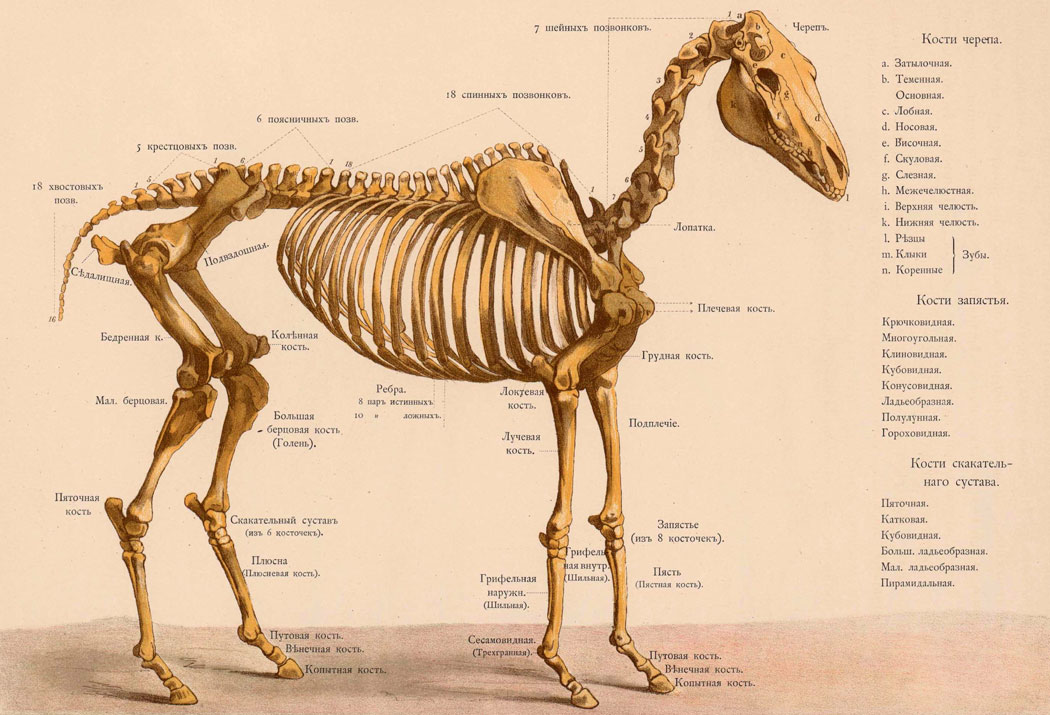
1.Филогенетический ряд лошади ( к слайду 6 )
Предком ствола современных лошадей считается живший в эоцене эогиппус. Эогиппус был высотой всего до 30 см, жил во влажных тропических лесах и, судя по строению зубов, питался семенами и плодами. При беге он опирался на все фаланги четырех пальцев (у его предков было еще пять пальцев) передней ноги и трех пальцев задней.
В слоях, отложившихся спустя несколько миллионов лет, находят эпигиппуса. Это небольшое лесное животное при беге опиралось уже лишь на последние фаланги четырех пальцев передней ноги.
Прошло еще около десятка миллионов лет, и эпигиппуса сменяет в находках мезогиппус: у него только три пальца на передней ноге, причем средний палец развит значительно сильнее боковых.
Еще через несколько миллионов лет мезогиппуса сменяет миогиппус, у которого были еще более короткие боковые пальцы и чуть большие размеры тела. Но все эти формы пока еще не похожи на лошадей, какими их знаем мы сегодня,— это небольшие лесные животные, не более крупной собаки. В результате изменения климата Земли сократилась площадь тропических лесов и расширилась зона саванн. Именно в этот период предки лошадей вышли из леса на открытые пространства саванн.
В горных породах, возраст которых на несколько миллионов лет меньше пород, вмещавших остатки миогиппуса, палеонтологи находят парагиппуса. Парагиппус жил небольшими стадами на открытых пространствах, при беге опирался лишь на средний палец, хотя второй и четвертый были еще заметно развиты: судя но строению зубов, питался лишь травой.
В плиоцене — периоде, отдаленном от нашего времени всего на 10 15 миллионов лет,— происходит дальнейшее расширение зоны степей и саванн. В это время возникают две ветви непарнокопытных: современные лошади и трехпалые гиппарионы. Гиппарионы, достигнув расцвета в своем развитии, полностью вымерли несколько миллионов лет назад.
Гиппарионы, достигнув расцвета в своем развитии, полностью вымерли несколько миллионов лет назад.
Непосредственный предок современных лошадей — плиогиппус, живший около двух миллионов лет назад. Сравнивая эогипнуса с современной лошадью, трудно установить их историческую, филогенетическую связь. Но если заполнить пространство между этими крайними звеньями многочисленными формами ископаемых форм копытных, живших в разное время в Северной Америке и Евразии, то возникает непрерывный филогенетический ряд. Подобные ряды сейчас известны для многих групп организмов.
Источник: http://pandia.ru/text/78/058/32567.php
2. Материалы из книги Ч. Дарвина
3. «Билеты»
Экскурсия: «ПРОЙДЁМ ПУТЁМ ЭВОЛЮЦИИ» I ОТДЕЛ: ПАЛЕОНТОЛОГИЧЕСКИЙ ПАЛЕОНТОЛОГИЯ- ………………………… …………………………………………………… ИСКОПАЕМЫЕ ФОРМЫ-………………………………………………………….. …………………………………………. ЭВОЛЮЦИОННЫЕ РЯДЫ- ………………………… II ОТДЕЛ: ЭМБРИОЛОГИЧЕСКИЙ ЭМБРИОН-……………………………………………… III ОТДЕЛ: СРАВНИТЕЛЬНО-АНАТОМИЧЕСКИЙ АНАТОМИЯ-……………………………………………. ………………………………………………………………. ГОМОЛОГИЧНЫЕ ОРГАНЫ-……………………………………………………… ……………………………………………………………… ……………………………………………………………… примеры:…………………………………………………… АНАЛОГИЧНЫЕ ОРГАНЫ-……………………………… ………………………………………………………………… примеры:……………………………………………………… РУДИМЕНТЫ-……………………………………………… ……………………………………………………………… примеры:…………………………………………………… АТАВИЗМЫ-……………………………………………… ……………………………………………………………… примеры:…………………………………………………… | ФАМИЛИЯ, ИМЯ………………………………………………. 1.КАКИЕ ДОКАЗАТЕЛЬСТВА ЭВОЛЮЦИИ ОСНОВЫВАЮТСЯ НА ДАННЫХ ОБ ИСКОПАЕМЫХ ОСТАНКАХ: А-МОРФОЛОГИЧЕСКИЕ Б- ЭМБРИОЛОГИЧЕСКИЕ В- ПАЛЕОНТОЛОГИЧЕСКИЕ Г-БИОГЕОГРАФИЧЕСКИЕ 2. А- КИСТЕПЁРАЯ РЫБА Б- УТКОНОС В- АРХЕОПТЕРИКС Г- ПРЫТКАЯ ЯЩЕРИЦА 3.КАКИЕ ОРГАНЫ ЛОШАДЕЙ ПРЕТЕРПЕВАЛИ НАИБОЛЬШИЕ ИЗМЕНЕНИЯ: А- КОНЕЧНОСТИ Б- СЕРДЦЕ В- РАЗМЕРЫ ТЕЛА Г- ЛЁГКИЕ 4. НАЗОВИТЕ ГОМОЛОГИЧНЫЕ ОРГАНЫ: А- ПЕРЕДНИЕ КОНЕЧНОСТИ ПОЗВОНОЧНЫХ Б- КРЫЛО БАБОЧКИ И КРЫЛО ПТИЦЫ В- ЧЕТВЁРТЫЙ ПАЛЕЦ КРЫЛА ПТИЦЫ Г-ТРЁХПАЛОСТЬ У ЛОШАДЕЙ 5. НАЗОВИТЕ АНАЛОГИЧНЫЕ ОРГАНЫ: А- ПЕРЕДНИЕ КОНЕЧНОСТИ ПОЗВОНОЧНЫХ Б- КРЫЛО БАБОЧКИ И КРЫЛО ПТИЦЫ В- ЧЕТВЁРТЫЙ ПАЛЕЦ КРЫЛА ПТИЦЫ Г- ТРЁХПАЛОСТЬ У ЛОШАДЕЙ 6.НАЗОВИТЕ РУДИМЕНТАРНЫЕ ОРГАНЫ: А- ПЕРЕДНИЕ КОНЕЧНОСТИ ПОЗВОНОЧНЫХ Б- КРЫЛО БАБОЧКИ И КРЫЛО ПТИЦЫ В- ЧЕТВЁРТЫЙ ПАЛЕЦ КРЫЛА ПТИЦЫ Г- ТРЁХПАЛОСТЬ У ЛОШАДЕЙ 7. А- ПЕРЕДНИЕ КОНЕЧНОСТИ ПОЗВОНОЧНЫХ Б- КРЫЛО БАБОЧКИ И КРЫЛО ПТИЦЫ В- ЧЕТВЁРТЫЙ ПАЛЕЦ КРЫЛА ПТИЦЫ Г- ТРЁХПАЛОСТЬ У ЛОШАДЕЙ 8. НА КАКИХ ЭТАПАХ РАЗВИТИЯ ЗАРОДЫШИ ПОЗВОНОЧНЫХ ОБНАРУЖИВАЮТ НАИБОЛЬШЕЕ СХОДСТВО МЕЖДУ СОБОЙ: А- НА РАННИХ ЭТАПАХ РАЗВИТИЯ Б- НА ПОЗДНИХ ЭТАПАХ РАЗВИТИЯ В- НА ПРОТЯЖЕНИИ ВСЕГО ПЕРИОДА РАЗВИТИЯ Г- НЕ ОБНАРУЖИВАЮТ СХОДСТВА |
 КАКИЕ ОРГАНИЗМЫ НЕ ЯВЛЯЮТСЯ ПЕРЕХОДНЫМИ ФОРМАМИ:
КАКИЕ ОРГАНИЗМЫ НЕ ЯВЛЯЮТСЯ ПЕРЕХОДНЫМИ ФОРМАМИ: НАЗОВИТЕ АТАВИЗМЫ:
НАЗОВИТЕ АТАВИЗМЫ:
Рост и развитие костей у лошади: когда скелет лошади становится зрелым?
1. Роджерс К.В., Болуэлл К.Ф., Джи Э.К., Розановский С.М. Развитие и производительность опорно-двигательного аппарата лошадей: влияние производственной системы и раннего обучения. Аним. Произв. науч. 2020;60:2069–2079. дои: 10.1071/AN17685. [CrossRef] [Google Scholar]
2. Rogers C.W., Dittmer K.E. Программируют ли детские игры скелетно-мышечную систему лошадей? Животные. 2019;9:646. doi: 10.3390/ani9090646. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
doi: 10.3390/ani9090646. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
3. Таннер Дж. К., Роджерс К. В., Ферт Э. С. Связь вех в обучении двухлетних детей с продолжительностью карьеры и успехами в скачках в выборке чистокровных лошадей в Новой Зеландии. Лошадиный ветеринар. Дж. 2013; 45:20–24. doi: 10.1111/j.2042-3306.2011.00534.x. [PubMed] [CrossRef] [Google Scholar]
4. Rogers C.W., Firth E.C., McIlwraith C.W., Barneveld A., Goodship AE, Kawcak C.E., Smith R.K.W., van Weeren P.R. Оценка новой стратегии модуляции развития скелета у скаковых лошадей путем навязывания беговых упражнений во время роста: влияние на 2- и 3-летнюю гоночную карьеру. Лошадиный ветеринар. Дж. 2008; 40:119–127. doi: 10.2746/042516408X266088. [PubMed] [CrossRef] [Google Scholar]
5. Rogers C.W., Firth E.C., McIlwraitth C.W., Barneveld A., Goodship A.E., Kawcak C.E., Smith R.K.W., van Weeren P.R. Оценка новой стратегии модуляции развития скелета чистокровных пород производительных лошадей, вводя упражнения на основе гусеницы во время роста.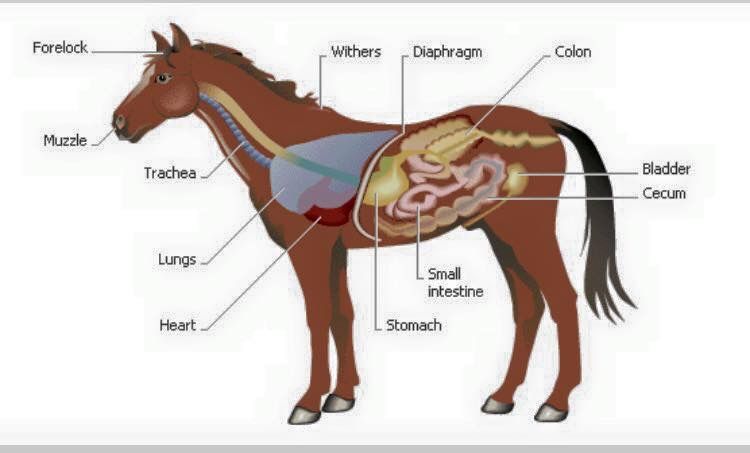 Лошадиный ветеринар. Дж. 2008; 40:111–118. doi: 10.2746/042516408X268923. [PubMed] [CrossRef] [Google Scholar]
Лошадиный ветеринар. Дж. 2008; 40:111–118. doi: 10.2746/042516408X268923. [PubMed] [CrossRef] [Google Scholar]
6. Вели Б.Д., Найт П.К., Томсон П.С., Уэйд С.М., Гамильтон Н.А. Связь возраста при первом старте с продолжительностью карьеры в популяции австралийских чистокровных скаковых лошадей. Лошадиный ветеринар. Дж. 2013; 45:410–413. doi: 10.1111/j.2042-3306.2012.00651.x. [PubMed] [CrossRef] [Академия Google]
7. Кочер А., Станиар В.Б. На характер роста чистокровных влияет дата рождения жеребят. Livest. науч. 2013; 154: 204–214. doi: 10.1016/j.livsci.2013.03.008. [CrossRef] [Google Scholar]
8. Sanders J.O., Qiu X., Lu X., Duren D.L., Liu R.W., Dang D., Menendez M.E., Hans S.D., Weber D.R., Cooperman D.R. Единая модель роста и созревания скелета во время скачка роста в подростковом возрасте. науч. 2017;7:16705. doi: 10.1038/s41598-017-16996-w. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
9. Хэмпсон Б.А., Поллит С.С. GPS-анализ активности домашних кобыл и новорождённых жеребят; Материалы 6-й Международной конференции по локомоции лошадей; Кабур, Франция. 16–19 июня 2008 г.; п. 17. [Google Scholar]
16–19 июня 2008 г.; п. 17. [Google Scholar]
10. Strand E., Braathen L.C., Hellsten M.C., Huse-Olsen L., Bjornsdottir S. Рентгенографическое время закрытия аппендикулярных пластинок роста у исландской лошади. Acta Veterinaria Scandinavica. 2007; 49:1–7. дои: 10.1186/1751-0147-49-19. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
11. Салхотра А., Шах Х.Н., Леви Б., Лонгакер М.Т. Механизмы развития и восстановления костей. Нац. Преподобный Мол. Клеточная биол. 2020; 21: 696–711. doi: 10.1038/s41580-020-00279-w. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
12. Craig L.E., Dittmer K.E., Thompson K.G. Кости и суставы. В: Макси Г., редактор. Джубб, Кеннеди и Палмер «Патология домашних животных». 6-е изд. Академическая пресса; Кембридж, Массачусетс, США: 2016. стр. 16–163. [Google Scholar]
13. Уильямс С., Алхатиб Б., Серра Р. Развитие осевого скелета и межпозвоночных дисков. Курс. Вершина. Дев. биол. 2019;133:49–90. [Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Прабху Л., Саралайя В., Пай М., Ранаде А., Сингх Г., Мадхьястха С. Целостность тела позвонка: обзор различных вовлеченных анатомических факторов в области поясницы. Остеопорос. Междунар. 2007; 18:891–903. [PubMed] [Google Scholar]
Прабху Л., Саралайя В., Пай М., Ранаде А., Сингх Г., Мадхьястха С. Целостность тела позвонка: обзор различных вовлеченных анатомических факторов в области поясницы. Остеопорос. Междунар. 2007; 18:891–903. [PubMed] [Google Scholar]
15. Knipe H., Chmiel-Nowak M. Центры окостенения позвоночного столба. Радиопедия. 2021 г.: 10.53347/rID-24695. [CrossRef] [Google Scholar]
16. Wojtków M., Głowacki M., Pezowicz C. Многомасштабная структурная характеристика замыкательной пластинки позвонка в моделях животных. Дж. Анат. 2021;239: 70–80. doi: 10.1111/joa.13402. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
17. Bick E.M., Copel J.W. Продольный рост человеческого позвонка: вклад в человеческую остегению. Дж. Боун Дж. Surg. 1950; 32: 803–814. doi: 10.2106/00004623-195032040-00008. [PubMed] [CrossRef] [Google Scholar]
18. Фрост Х.М. Костная масса и механостат — предложение. Анат. Рек. 1987; 219:1–9. doi: 10.1002/ar.10921
. [PubMed] [CrossRef] [Google Scholar]
19. Фрост Х.М. Парадигма скелетной физиологии штата Юта: обзор ее идей в отношении костей, хрящей и органов из коллагеновой ткани. Дж. Боун Шахтер. Метаб. 2000;18:305–316. doi: 10.1007/s007740070001. [PubMed] [CrossRef] [Академия Google]
Фрост Х.М. Парадигма скелетной физиологии штата Юта: обзор ее идей в отношении костей, хрящей и органов из коллагеновой ткани. Дж. Боун Шахтер. Метаб. 2000;18:305–316. doi: 10.1007/s007740070001. [PubMed] [CrossRef] [Академия Google]
20. Маткович В., Елич Т., Уордлоу Г.М., Ильич Дж.З., Гоэль П.К., Райт Дж.К., Андон М.Б., Смит К.Т., Хини Р.П. Время пиковой костной массы у женщин европеоидной расы и ее значение для профилактики остеопороза. Вывод из модели поперечного сечения. Дж. Клин. расследование 1994; 93: 799–808. doi: 10.1172/JCI117034. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
21. Бек-Нильсен С.С., Греджио Н.А., Хагенас Л. Определение растущего и созревающего скелета и его значение при заболеваниях, влияющих на рост скелета, таких как X- Связанная гипофосфатемия (XLH) Int. Дж. Редкий Дис. Беспорядок. 2021;4:13. дои: 10.23937/2643-4571/1710029. [CrossRef] [Google Scholar]
22. Hawkes C.P., Mostoufi-Moab S., McCormack S.E., Grimberg A., Zemel B. S. Справочные таблицы отношения роста сидя к росту стоя для детей в Соединенных Штатах. Дж. Педиатрия. 2020;226:221–227.e15. doi: 10.1016/j.jpeds.2020.06.051. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
S. Справочные таблицы отношения роста сидя к росту стоя для детей в Соединенных Штатах. Дж. Педиатрия. 2020;226:221–227.e15. doi: 10.1016/j.jpeds.2020.06.051. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
23. Даллора А.Л., Андерберг П., Квист О., Мендес Э., Диас Руис С., Санмартин Берглунд Дж. Оценка костного возраста с помощью различных методов машинного обучения методы: систематический обзор литературы и метаанализ. ПЛОС ОДИН. 2019;14:e0220242. doi: 10.1371/journal.pone.0220242. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
24. Кэмерон Н. Можно ли использовать показатели зрелости для оценки хронологического возраста детей? Анна. Гум. биол. 2015;42:302–307. doi: 10.3109/03014460.2015.1032349. [PubMed] [CrossRef] [Google Scholar]
25. Фредрикс А. М., ван Бюрен С., Ван Хил В., Дейкман-Ниринкс Р., Верлоове-Ванхорик С., Вит Дж. М. Общенациональные возрастные нормы роста сидя и длины ног , соотношение роста и роста в сидячем положении, а также их диагностическое значение при диспропорциональных нарушениях роста. Арка Дис. Ребенок. 2005;90: 807–812. doi: 10.1136/adc.2004.050799. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Арка Дис. Ребенок. 2005;90: 807–812. doi: 10.1136/adc.2004.050799. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
26. Хантингтон П.Дж., Браун-Дуглас К.Г., Пэган Дж.Д. Рост и развитие чистокровных лошадей. Аним. Произв. науч. 2020;60:2093–2102. дои: 10.1071/AN19629. [CrossRef] [Google Scholar]
27. Brown-Douglas C.G., Firth E.C., Parkinson T.J., Fennessy P.F. Начало половой зрелости у чистокровных лошадей, выращиваемых на пастбищах, рожденных в южном полушарии весной и осенью. Лошадиный ветеринар. Дж. 2004; 36: 499–504. doi: 10.2746/0425164044877422. [PubMed] [CrossRef] [Академия Google]
28. Van Weeren P., Sloet van Oldruitenborgh-Oosterbaan M., Barneveld A. Влияние массы тела при рождении, темпов прибавки массы тела и конечного достигнутого роста и пола на развитие остеохондротических поражений в популяции генетически предрасположенных теплокровных жеребят. . Лошадиный ветеринар. Дж. 1999; 31: 26–30. doi: 10.1111/j.2042-3306.1999.tb05310.x. [PubMed] [CrossRef] [Google Scholar]
29. Валетт Дж. П., Роберт С., Денуа Дж. М. Использование линейных и нелинейных функций для описания роста молодых спортивных и скаковых лошадей, родившихся в Нормандии. Животное. 2008; 2: 560–565. doi: 10.1017/S1751731107001462. [PubMed] [CrossRef] [Академия Google]
Валетт Дж. П., Роберт С., Денуа Дж. М. Использование линейных и нелинейных функций для описания роста молодых спортивных и скаковых лошадей, родившихся в Нормандии. Животное. 2008; 2: 560–565. doi: 10.1017/S1751731107001462. [PubMed] [CrossRef] [Академия Google]
30. Peugnet P., Mendoza L., Wimel L., Duchamp G., Dubois C., Reigner F., Caudron I., Deliège B., Toquet M.-P., Richard E. Продольное исследование роста и костно-суставной статус у жеребят, рожденных в результате межпородного переноса эмбрионов. Дж. Ветеринар по лошадям. науч. 2016; 37:24–38. doi: 10.1016/j.jevs.2015.11.008. [CrossRef] [Google Scholar]
31. Эллис Р.Н.В., Лоуренс Т.Л.Дж. Энергетическая недостаточность у отъемыша кобылки. 2. Влияние на конформацию тела и закрытие эпифизарной пластинки на передней конечности. бр. Вет. Дж. 1978;134:322–332. doi: 10.1016/S0007-1935(17)33434-6. [PubMed] [CrossRef] [Google Scholar]
32. Franco D., Crecente S., Vázquez J.A., Gómez M., Lorenzo J.M. Влияние скрещивания и количества откормочного рациона на параметры роста, туши и мясной состав жеребят забит в возрасте 15 мес. Мясная наука. 2013;93:547–556. doi: 10.1016/j.meatsci.2012.11.018. [PubMed] [CrossRef] [Google Scholar]
Мясная наука. 2013;93:547–556. doi: 10.1016/j.meatsci.2012.11.018. [PubMed] [CrossRef] [Google Scholar]
33. McManus C.M., Louvandini H., Campos V.A.L. Нелинейные кривые роста веса и роста в четырех генетических группах лошадей. Сиенсия Аним. Бюстгальтеры. 2010;11 [Google Академия]
34. Ловшин Э., Фазаринц Г., Погачник А., Бавдек С.В. Динамика роста липицанских лошадей и их сравнение с другими породами лошадей. Арка Пфлюгера. 2001;442:r211–r212. doi: 10.1007/s004240100029. [PubMed] [CrossRef] [Google Scholar]
35. Veraa S., Scheffer CJ, Smeets D.H., de Bruin R.B., Hoogendoorn A.C., Vernooij J.C., Nielen M., Back W. Шейный индекс ширины диска является надежным параметром и характерен для молодых растущих голландских теплокровных лошадей. Вет. Радиол. УЗИ. 2021; 62: 11–19. doi: 10.1111/vru.12913. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
36. Hasler C., Sprecher C.M., Milz S. Сравнение позвоночника незрелой овцы и растущего позвоночника человека: спондилометрическая база данных для исследования модуляции роста. Позвоночник. 2010; 35:E1262–E1272. doi: 10.1097/BRS.0b013e3181e24196. [PubMed] [CrossRef] [Google Scholar]
Позвоночник. 2010; 35:E1262–E1272. doi: 10.1097/BRS.0b013e3181e24196. [PubMed] [CrossRef] [Google Scholar]
37. Томпсон К.Н. Темпы роста скелета отъемышей и годовалых чистокровных лошадей. Дж. Аним. науч. 1995; 73: 2513–2517. дои: 10.2527/1995,7392513х. [PubMed] [CrossRef] [Google Scholar]
38. Батлер Дж.А., Коллес С.М., Дайсон С., Колд С., Поулос П. Клиническая радиология лошади. Блэквелл Сайентифик Публикейшнз Лтд.; Oxford, UK: 1993. [Google Scholar]
39. Firth E.C., Rogers C.W., van Weeren P.R., Barneveld A., McIlwraith C.W., Kawcak C.E., Goodship A.E., Smith R.K.W. Влияние предыдущих кондиционных упражнений на реакции диафизарных и метафизарных костей на введение и отмену тренировок у молодых чистокровных лошадей. Вет. Дж. 2012; 192:34–40. doi: 10.1016/j.tvjl.2011.05.016. [PubMed] [CrossRef] [Google Scholar]
40. Fretz P.B., Cymbaluk N.F., Pharr J.W. Количественный анализ роста длинных костей у лошади. Являюсь. Дж. Вет. Рез. 1984; 45: 1602–1609. [PubMed] [Google Scholar]
41. Вулкано Л.С., Мамприм М.Дж., Муниз Л.М., Морейра А.Ф., Луна С.П. Рентгенографическое исследование закрытия дистального лучевого физа у чистокровных лошадей. Вет. Радиол. УЗИ. 1997; 38: 352–354. doi: 10.1111/j.1740-8261.1997.tb02096.x. [PubMed] [CrossRef] [Академия Google]
Вулкано Л.С., Мамприм М.Дж., Муниз Л.М., Морейра А.Ф., Луна С.П. Рентгенографическое исследование закрытия дистального лучевого физа у чистокровных лошадей. Вет. Радиол. УЗИ. 1997; 38: 352–354. doi: 10.1111/j.1740-8261.1997.tb02096.x. [PubMed] [CrossRef] [Академия Google]
42. Yoshida K., Ueda Y., Masumitsu H. Рентгенологические исследования окостенения у чистокровных 2. Замыкательный процесс в дистальных эпифизарных линиях лучевой кости и 3-й пястной кости и проксимальной эпифизарной линии проксимальной фаланги, и система оценки зрелости костей. Бык. Конный Рез. Инст. 1982; 1982: 18–29. [Google Scholar]
43. Бэнкс В.К. Рентгенография костного мозга и ее использование в дрессировке чистокровных лошадей. Акта Радиол. Диагн. 1972; 12: 71–72. дои: 10.3109/05678067209173996. [PubMed] [CrossRef] [Google Scholar]
44. Łuszczyński J., Pieszka M., Kosiniak-Kamysz K. Влияние породы и пола лошади на скорость роста и рентгенографическое время закрытия дистальной радиальной пластинки роста метафиза.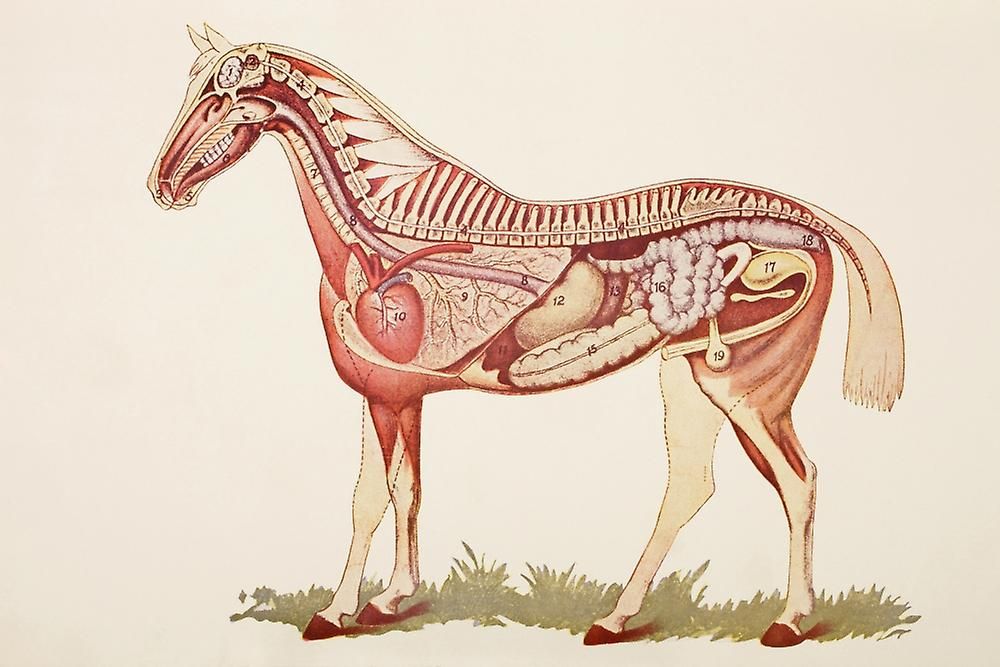 Livest. науч. 2011; 141: 252–258. doi: 10.1016/j.livsci.2011.06.009. [CrossRef] [Google Scholar]
Livest. науч. 2011; 141: 252–258. doi: 10.1016/j.livsci.2011.06.009. [CrossRef] [Google Scholar]
45. Gabel A., Spencer C., Pipers F. Изучение корреляции закрытия дистального отдела лучевой кости с производительностью и травмами в Standardbred. Варенье. Вет. Мед. доц. 1977;170:188–194. [PubMed] [Google Scholar]
46. Ульхорн Х., Экселл П., Карлстен Дж. Сцинтиграфическая характеристика закрытия дистального лучевого физиоза у молодых скаковых лошадей стандартной породы. Вет. Радиол. УЗИ. 2000;41:181–186. doi: 10.1111/j.1740-8261.2000.tb01475.x. [PubMed] [CrossRef] [Google Scholar]
47. Pezzoli G., del Bue M. Valuatazione radiografica del grado di sviluppo scheletrico nel cavallo trottatore in rapporto all’attivita atletica. Фолиа Вет. лат. 1975; 5: 399–411. [PubMed] [Академия Google]
48. Пазолини М., Меомартино Л., Теста А., Фатоне Г., Потена А., Роза Г.Д., Ламанья Ф. Рентгенологическая оценка зрелости скелета у скаковой лошади: статистическая проверка и корреляция с ортопедическими травмами в стандартной породе. Иппология. 2007; 18:15–19. [Google Scholar]
Иппология. 2007; 18:15–19. [Google Scholar]
49. Майерс В., мл., Эммерсон М. Возраст и характер закрытия эпифизов на передних конечностях двух арабских жеребят 1. Ветеринар. Радиол. 1966; 7: 39–47. doi: 10.1111/j.1740-8261.1966.tb01062.x. [Перекрестная ссылка] [Академия Google]
50. Mamprim M., Vulcano L., Muniz L. Радиографическое исследование дистального эпифиза дистального отдела лучевой кости Manga-Larga (Рентгенографическое исследование эпифизарного закрытия дистального отдела лучевой кости у кобылок Manga-Larga.) Vet. Зут. 1992; 4: 59–62. [Google Scholar]
51. Коскинен Э., Катила Т. Влияние 19-норандростенололилаурата на концентрацию тестостерона в сыворотке, либидо и закрытие дистальной радиальной зоны роста у жеребят. Акта Вет. Сканд. 1997; 38: 59–67. doi: 10.1186/BF03548508. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
52. Тухман С., Таю М., Шульц Дж., Земель Б.С., Бернхэм Дж.М., Леонард М.Б. Интерпретация биомаркеров костного метаболизма у детей: влияние скорости роста и размеров тела у здоровых детей и при хронических заболеваниях. Дж. Педиатрия. 2008;153:484–490.e2. doi: 10.1016/j.jpeds.2008.04.028. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Дж. Педиатрия. 2008;153:484–490.e2. doi: 10.1016/j.jpeds.2008.04.028. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
53. Monjardino T., Silva P., Amaro J., Carvalho O., Guimarães J.T., Santos A.C., Lucas R. Маркеры формирования кости и резорбции в 7 лет: связь с ростом и минерализацией костей. ПЛОС ОДИН. 2019;14:e0219423. doi: 10.1371/journal.pone.0219423. [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
54. Кадоган Дж., Блумсон А., Баркер М.Е., Истелл Р. Продольное исследование увеличения костной массы у девочек полового созревания: антропометрические и биохимические корреляты. Дж. Боун Шахтер. Рез. 1998; 13:1602–1612. doi: 10.1359/jbmr.1998.13.10.1602. [PubMed] [CrossRef] [Google Scholar]
55. Terpstra L., Knol D., Van Coeverden S., Delemarre-van de Waal H. Маркеры костного метаболизма предсказывают увеличение костной массы, роста и высоты сидя во время полового созревания в зависимости от на генотип VDR Fok1. клин. Эндокринол. 2006; 64: 625–631. doi: 10.1111/j.1365-2265.2006.02516.x. [PubMed] [CrossRef] [Академия Google]
doi: 10.1111/j.1365-2265.2006.02516.x. [PubMed] [CrossRef] [Академия Google]
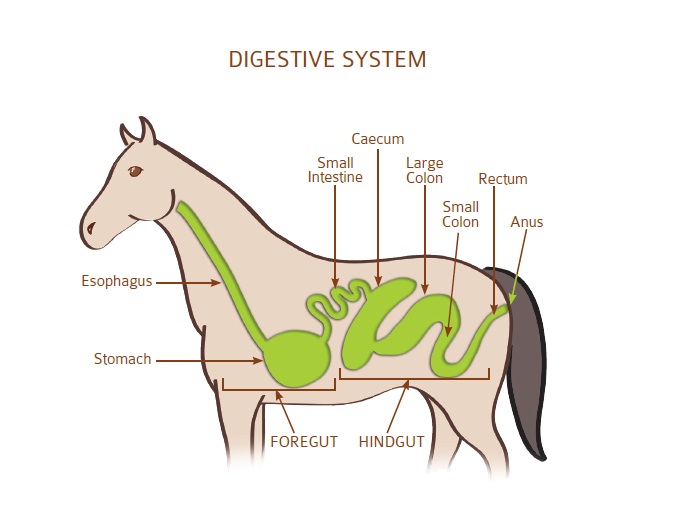
Пищеварительная система лошади | HyGain Horse Feed – Hygain US
Командный маркетинг | 18.10.20
Воспроизведено с разрешения QA International из книги The Visual Dictionary. © QA International, 2003. Все права защищены.
Лошадь — нежвачное травоядное. Нежвачный означает, что у лошадей нет многокамерных желудков, как у крупного рогатого скота. Вместо этого у лошади простой желудок, который работает так же, как у человека. Травоядные означает, что лошади живут на диете из растительного материала. Пищеварительный тракт лошадей уникален тем, что он ферментативно переваривает порции корма сначала в передней кишке, а ферментирует в задней кишке. Пищеварительную систему лошади действительно следует рассматривать как состоящую из двух частей. Первый отдел имеет сходство с прецекальной пищеварительной системой животных с однокамерным желудком, таких как собака, человек или свинья. Второй отдел больше похож на рубец коровы. Это сильно влияет на то, как мы должны думать о кормлении лошадей, находящихся под нашим присмотром. Однако лошадь не является ни собакой, ни жвачным животным, ни даже прямой комбинацией того и другого. Он уникален и должен рассматриваться как таковой. Корова выигрывает от микробного расщепления волокнистой пищи в начале ЖКТ (желудочно-кишечного тракта), а всасывание питательных веществ может происходить по всему кишечнику. Пищевой белок не используется эффективно, потому что микробная ферментация расщепляет белок и некоторое количество углеводов. У лошади, в отличие от жвачных, микробное брожение происходит после «моногастрального» раздела, а не до него. Это оказывает большое влияние на то, как мы должны кормить лошадь, и частично объясняет, почему лошадь и корова так сильно различаются по эффективности и потребностям в питании.
Второй отдел больше похож на рубец коровы. Это сильно влияет на то, как мы должны думать о кормлении лошадей, находящихся под нашим присмотром. Однако лошадь не является ни собакой, ни жвачным животным, ни даже прямой комбинацией того и другого. Он уникален и должен рассматриваться как таковой. Корова выигрывает от микробного расщепления волокнистой пищи в начале ЖКТ (желудочно-кишечного тракта), а всасывание питательных веществ может происходить по всему кишечнику. Пищевой белок не используется эффективно, потому что микробная ферментация расщепляет белок и некоторое количество углеводов. У лошади, в отличие от жвачных, микробное брожение происходит после «моногастрального» раздела, а не до него. Это оказывает большое влияние на то, как мы должны кормить лошадь, и частично объясняет, почему лошадь и корова так сильно различаются по эффективности и потребностям в питании.
Рот
Лошади захватывают пищу, используя комбинацию губ, языка и зубов. Губы лошадей чрезвычайно осязательны, когда дело доходит до потребления корма. Они могут быть довольно избирательными, так как многие из нас видели бы порошкообразные добавки или гранулы в красивой небольшой кучке у бутылочки с кормушкой. Корм смешивается со слюной во рту, образуя влажный болюс, который можно легко проглотить. Три пары желез вырабатывают слюну — околоушные, подчелюстные и подъязычные. Лошади производят от 20 до 80 литров слюны в день. Шалфей содержит бикарбонат, который буферизует и защищает аминокислоты в высококислотном желудке. Слюна также содержит небольшое количество амилазы, которая способствует перевариванию углеводов. Во рту 36 зубов (самки) и 40 зубов (самцы). Зубы волка не включены, так как они есть не у всех лошадей. Верхняя челюсть лошади шире нижней, что позволяет совершать довольно сложные жевательные движения. Жевательное действие лошади — это размашистое действие, которое включает в себя как боковые движения вперед и назад, так и вертикальные движения. Это позволяет эффективно измельчать корм и смешивать его со слюной, чтобы инициировать процесс пищеварения.
Они могут быть довольно избирательными, так как многие из нас видели бы порошкообразные добавки или гранулы в красивой небольшой кучке у бутылочки с кормушкой. Корм смешивается со слюной во рту, образуя влажный болюс, который можно легко проглотить. Три пары желез вырабатывают слюну — околоушные, подчелюстные и подъязычные. Лошади производят от 20 до 80 литров слюны в день. Шалфей содержит бикарбонат, который буферизует и защищает аминокислоты в высококислотном желудке. Слюна также содержит небольшое количество амилазы, которая способствует перевариванию углеводов. Во рту 36 зубов (самки) и 40 зубов (самцы). Зубы волка не включены, так как они есть не у всех лошадей. Верхняя челюсть лошади шире нижней, что позволяет совершать довольно сложные жевательные движения. Жевательное действие лошади — это размашистое действие, которое включает в себя как боковые движения вперед и назад, так и вертикальные движения. Это позволяет эффективно измельчать корм и смешивать его со слюной, чтобы инициировать процесс пищеварения. Текстура корма, скармливаемого лошадям, сильно влияет на скорость жевания (размахивания челюстями) и скорость приема пищи. Средняя лошадь с общими показателями совершает 60 000 взмахов челюстью в день при выпасе. Это количество будет резко уменьшено, если они ограничены конюшней и скармливают большое количество зерна. Размер лошади также влияет на время и количество взмахов челюстью, необходимых для достаточного пережевывания корма. Среднестатистической лошади весом 500 кг обычно требуется 40 минут и 3400 взмахов челюстью, чтобы съесть один килограмм сена. Пони обычно потребляют такое количество сена в два раза дольше. Овес, с другой стороны, занимает всего 10 минут и 850 взмахов челюстью для взрослой лошади и до пяти раз дольше для пони. Когда лошади жуют волокнистые корма, такие как сено или пастбище, челюсти тянутся длинными движениями. Вот почему у лошадей, постоянно находящихся на пастбищах, редко появляются острые края зубов. Зерна потребляются более коротким движением, которое не выходит за пределы внешнего края зубов.
Текстура корма, скармливаемого лошадям, сильно влияет на скорость жевания (размахивания челюстями) и скорость приема пищи. Средняя лошадь с общими показателями совершает 60 000 взмахов челюстью в день при выпасе. Это количество будет резко уменьшено, если они ограничены конюшней и скармливают большое количество зерна. Размер лошади также влияет на время и количество взмахов челюстью, необходимых для достаточного пережевывания корма. Среднестатистической лошади весом 500 кг обычно требуется 40 минут и 3400 взмахов челюстью, чтобы съесть один килограмм сена. Пони обычно потребляют такое количество сена в два раза дольше. Овес, с другой стороны, занимает всего 10 минут и 850 взмахов челюстью для взрослой лошади и до пяти раз дольше для пони. Когда лошади жуют волокнистые корма, такие как сено или пастбище, челюсти тянутся длинными движениями. Вот почему у лошадей, постоянно находящихся на пастбищах, редко появляются острые края зубов. Зерна потребляются более коротким движением, которое не выходит за пределы внешнего края зубов. Когда скармливается большое количество зерна, жевательная деятельность лошадей изменяется, и зубы изнашиваются неравномерно. Крючки или острые края начнут формироваться на внешнем крае зубов. Если зубы неправильно «плавают» или не скрежещут, скорость приема пищи, жевательная эффективность, аппетит и темперамент могут серьезно пострадать. Если корм не пережевывается должным образом, болюс (корм и шалфей) может застрять в пищеводе и вызвать удушье.
Когда скармливается большое количество зерна, жевательная деятельность лошадей изменяется, и зубы изнашиваются неравномерно. Крючки или острые края начнут формироваться на внешнем крае зубов. Если зубы неправильно «плавают» или не скрежещут, скорость приема пищи, жевательная эффективность, аппетит и темперамент могут серьезно пострадать. Если корм не пережевывается должным образом, болюс (корм и шалфей) может застрять в пищеводе и вызвать удушье.
Пищевод
Это простая мышечная трубка, по которой пища проходит изо рта в желудок. Длина пищевода у взрослой лошади составляет около 1,5 м. Поскольку пищевод довольно длинный, а рефлюксная способность лошади очень мала, неправильно пережеванные крупные куски корма, такие как морковь и т. д., могут застрять в пищеводе лошади и вызвать удушье. Вот почему важно правильно ухаживать за зубами лошадей, чтобы лошади достаточно пережевывали корм, а также не позволяли лошадям «засовывать» корм вниз, не пережевывая корм. Добавление мякины в корм для лошадей или помещение кирпича или большого камня в кормушку для лошадей снизит скорость потребления лошади и снизит риск удушья из-за того, что лошадь «бросает» свой корм.
Добавление мякины в корм для лошадей или помещение кирпича или большого камня в кормушку для лошадей снизит скорость потребления лошади и снизит риск удушья из-за того, что лошадь «бросает» свой корм.
Желудок
Желудок лошади небольшой по отношению к размерам животного и составляет всего 10 % вместимости пищеварительной системы или 9-15 литров в объеме. Естественная пищевая привычка лошади — часто есть небольшое количество грубых кормов. Одомашнивание внесло изменения во все это. В настоящее время ожидается, что лошади будут съедать большое количество зернового корма один или два раза в день, чтобы соответствовать нашему образу жизни. Это сильно подрывает пищеварительные способности и здоровье лошади. Было установлено, что мы можем улучшить пищеварительную эффективность лошади, часто кормя небольшими порциями (ассимилируя естественный выпас), но это было взвешено с учетом трудозатрат на это. В желудке корм смешивается с пепсином (фермент для переваривания белков) и соляной кислотой, помогающей расщеплять твердые частицы. Скорость прохождения корма через желудок сильно варьируется в зависимости от того, как кормят лошадь. Время прохождения может составлять всего 15 минут, когда лошадь ест много. Если лошадь голодает, ей потребуется 24 часа, чтобы желудок очистился. Давно встал вопрос, чем кормить лошадь в первую очередь, зерном или сеном. Из-за своей плотности зерна, как правило, дольше остаются в желудке, но не было доказано, что полезно кормить их в первую очередь. Для тех, кто быстро ест, в корм можно добавлять мякину, чтобы увеличить объем корма, что замедляет скорость потребления. Другой вопрос, должна ли лошадь получать воду до или после еды. Если вы оставите это на усмотрение лошади, она обычно будет пить немного во время еды, если потребляет сухие корма. Лучшая рекомендация — постоянно предлагать свежую чистую воду. Желудок имеет 3 основные области; слепой мешок, фундальная и пилорическая области. Каждый из них уникален по структуре и функциям. Область saccus caecus расположена у входа в желудок и пищевод.
Скорость прохождения корма через желудок сильно варьируется в зависимости от того, как кормят лошадь. Время прохождения может составлять всего 15 минут, когда лошадь ест много. Если лошадь голодает, ей потребуется 24 часа, чтобы желудок очистился. Давно встал вопрос, чем кормить лошадь в первую очередь, зерном или сеном. Из-за своей плотности зерна, как правило, дольше остаются в желудке, но не было доказано, что полезно кормить их в первую очередь. Для тех, кто быстро ест, в корм можно добавлять мякину, чтобы увеличить объем корма, что замедляет скорость потребления. Другой вопрос, должна ли лошадь получать воду до или после еды. Если вы оставите это на усмотрение лошади, она обычно будет пить немного во время еды, если потребляет сухие корма. Лучшая рекомендация — постоянно предлагать свежую чистую воду. Желудок имеет 3 основные области; слепой мешок, фундальная и пилорическая области. Каждый из них уникален по структуре и функциям. Область saccus caecus расположена у входа в желудок и пищевод. Когда пища попадает в желудок, она начинает попадать под влияние соляной кислоты и пепсина — фермента, переваривающего белок. Однако этот корм (особенно если это преимущественно трава) уже выделяет растворимые сахара для усвоения и подвергается бактериальной ферментации с образованием молочной кислоты. В нормальных условиях, когда соляная кислота смешивается с желудком, рН падает, ферментация замедляется и в конце концов прекращается. Это важный процесс, потому что, если этого не происходит, а брожение продолжается, относительно нерастяжимый желудок фиксированного объема очень быстро наполняется газом и, при небольшой возможности сброса давления через пищевод, может привести к желудочным коликам или к экстремальным состояниям. случаи разрыва слизистой оболочки желудка. По мере продвижения корма через желудок следующим отделом желудка является фундальная область. Уровень pH снижается примерно до 5,4, и ферментация начинает приостанавливаться. Пепсин и желудочная кислота инициируют переваривание и расщепление липидов (жиров) и белков (аминокислот).
Когда пища попадает в желудок, она начинает попадать под влияние соляной кислоты и пепсина — фермента, переваривающего белок. Однако этот корм (особенно если это преимущественно трава) уже выделяет растворимые сахара для усвоения и подвергается бактериальной ферментации с образованием молочной кислоты. В нормальных условиях, когда соляная кислота смешивается с желудком, рН падает, ферментация замедляется и в конце концов прекращается. Это важный процесс, потому что, если этого не происходит, а брожение продолжается, относительно нерастяжимый желудок фиксированного объема очень быстро наполняется газом и, при небольшой возможности сброса давления через пищевод, может привести к желудочным коликам или к экстремальным состояниям. случаи разрыва слизистой оболочки желудка. По мере продвижения корма через желудок следующим отделом желудка является фундальная область. Уровень pH снижается примерно до 5,4, и ферментация начинает приостанавливаться. Пепсин и желудочная кислота инициируют переваривание и расщепление липидов (жиров) и белков (аминокислот). Конечным отделом желудка является пилорический отдел, где желудок соединяется с тонкой кишкой. Уровень pH снижается до 2,6, что практически уничтожает все способные к брожению молочнокислые бактерии. Протеолитическая активность (переваривание белков) в этой области в 15-20 раз выше, чем в фундальной области. Измененная практика кормления привела к тому, что в течение дня желудок лошадей практически пуст. Смесь корма и слюны смешивается с кислотой, вырабатываемой желудком. Когда желудок лошади пуст, кислота разрушает незащищенные чешуйчатые клетки области слепого мешка желудка. Это приводит к изъязвлению слизистой оболочки желудка. Исследования показали, что более 80% чистокровных имеют язву желудка в той или иной степени. Язвы желудка могут повлиять на аппетит, поведение и производительность лошади. Кормление лошадей более высокой долей грубых кормов в их рационе, небольшими частыми приемами пищи и предоставлением им возможности пастись резко снизит частоту и тяжесть язвы желудка.
Конечным отделом желудка является пилорический отдел, где желудок соединяется с тонкой кишкой. Уровень pH снижается до 2,6, что практически уничтожает все способные к брожению молочнокислые бактерии. Протеолитическая активность (переваривание белков) в этой области в 15-20 раз выше, чем в фундальной области. Измененная практика кормления привела к тому, что в течение дня желудок лошадей практически пуст. Смесь корма и слюны смешивается с кислотой, вырабатываемой желудком. Когда желудок лошади пуст, кислота разрушает незащищенные чешуйчатые клетки области слепого мешка желудка. Это приводит к изъязвлению слизистой оболочки желудка. Исследования показали, что более 80% чистокровных имеют язву желудка в той или иной степени. Язвы желудка могут повлиять на аппетит, поведение и производительность лошади. Кормление лошадей более высокой долей грубых кормов в их рационе, небольшими частыми приемами пищи и предоставлением им возможности пастись резко снизит частоту и тяжесть язвы желудка.
Тонкий кишечник
Дигестер переходит из желудка в тонкую кишку. Тонкий кишечник составляет примерно 28% пищеварительного тракта лошади, имеет длину 15-22 м и объем 55-70 литров. Это основное место пищеварения у современной спортивной лошади. Тонкая кишка разделена на 3 отдела; двенадцатиперстной, тощей и подвздошной кишках. Слюна лошади содержит лишь небольшое количество амилазы, и у большинства лошадей в желудке практически не происходит пищеварения. Таким образом, большая часть пищеварения происходит в тонком и толстом кишечнике. Хотя сам кишечник выделяет некоторые ферменты, поджелудочная железа выделяет их гораздо больше. В тонкой кишке процессы пищеварения (ферментативное расщепление белков, жиров, крахмалов и сахаров) сходны с таковыми у других животных с однокамерным желудком, но активность некоторых ферментов химуса (пищевой смеси), в частности амилазы, ниже, чем у животных с однокамерным желудком. у других моногастричных животных. В этом пищеварительном процессе много компонентов. Ферменты поджелудочной железы помогут переварить пищу; углеводы переваривают сахара и крахмалы; протеазы расщепляют белки до аминокислот; липазы и желчь из печени добавляются для эмульгирования (разбивания на более мелкие части) жиров и суспендирования жира в воде.
Тонкий кишечник составляет примерно 28% пищеварительного тракта лошади, имеет длину 15-22 м и объем 55-70 литров. Это основное место пищеварения у современной спортивной лошади. Тонкая кишка разделена на 3 отдела; двенадцатиперстной, тощей и подвздошной кишках. Слюна лошади содержит лишь небольшое количество амилазы, и у большинства лошадей в желудке практически не происходит пищеварения. Таким образом, большая часть пищеварения происходит в тонком и толстом кишечнике. Хотя сам кишечник выделяет некоторые ферменты, поджелудочная железа выделяет их гораздо больше. В тонкой кишке процессы пищеварения (ферментативное расщепление белков, жиров, крахмалов и сахаров) сходны с таковыми у других животных с однокамерным желудком, но активность некоторых ферментов химуса (пищевой смеси), в частности амилазы, ниже, чем у животных с однокамерным желудком. у других моногастричных животных. В этом пищеварительном процессе много компонентов. Ферменты поджелудочной железы помогут переварить пищу; углеводы переваривают сахара и крахмалы; протеазы расщепляют белки до аминокислот; липазы и желчь из печени добавляются для эмульгирования (разбивания на более мелкие части) жиров и суспендирования жира в воде. Желчь постоянно поступает в тонкую кишку из печени, потому что у лошади нет желчного пузыря для ее хранения. Сок поджелудочной железы также содержит некоторое количество щелочи и бикарбонатов, которые буферизуют кислоту, поступающую из желудка (кормовой болюс), покидающую желудок, и помогают поддерживать оптимальную среду для функционирования пищеварительных ферментов. После переваривания пищи она всасывается через стенки тонкой кишки и с током крови разносится по клеткам, нуждающимся в питательных веществах. Почти 30-60% переваривания и всасывания углеводов и почти всего всасывания аминокислот происходит в тонком кишечнике. В тонком кишечнике всасываются жирорастворимые витамины A, D, E и K, а также некоторые минералы, такие как кальций и фосфор. Изменение структуры углеводов корма с помощью таких процессов, как микронизация, значительно повышает переваримость зерна в тонком кишечнике примерно до 90%. Это снижает нагрузку на толстую кишку и может снизить риск перегрузки пищеварительного тракта и возникновения колик, ламинита и ацидоза.
Желчь постоянно поступает в тонкую кишку из печени, потому что у лошади нет желчного пузыря для ее хранения. Сок поджелудочной железы также содержит некоторое количество щелочи и бикарбонатов, которые буферизуют кислоту, поступающую из желудка (кормовой болюс), покидающую желудок, и помогают поддерживать оптимальную среду для функционирования пищеварительных ферментов. После переваривания пищи она всасывается через стенки тонкой кишки и с током крови разносится по клеткам, нуждающимся в питательных веществах. Почти 30-60% переваривания и всасывания углеводов и почти всего всасывания аминокислот происходит в тонком кишечнике. В тонком кишечнике всасываются жирорастворимые витамины A, D, E и K, а также некоторые минералы, такие как кальций и фосфор. Изменение структуры углеводов корма с помощью таких процессов, как микронизация, значительно повышает переваримость зерна в тонком кишечнике примерно до 90%. Это снижает нагрузку на толстую кишку и может снизить риск перегрузки пищеварительного тракта и возникновения колик, ламинита и ацидоза. Прохождение пищи через тонкую кишку может занять от 30 до 60 минут, так как большая часть пищеварительного тракта движется со скоростью примерно 30 см в минуту. Однако корм обычно проходит через тонкий кишечник за 3-4 часа. Чем быстрее дигеста проходит через тонкий кишечник, тем меньше времени остается на действие ферментов. Было показано, что добавление масла в рацион лошадей уменьшает поток корма через тонкую кишку, что дает пищеварительным ферментам больше времени для переработки крахмалов, белков и жиров, тем самым увеличивая общую усвояемость этих питательных веществ трактом и максимально улучшая пищеварение тонкого кишечника. эффективность. Лошади очень восприимчивы к коликам или смерти от токсичных материалов в корме. В отличие от коровы, у которой в рубце есть бактерии, которые могут обезвреживать материалы до того, как они достигнут тонкой кишки, токсичные материалы, которые лошадь может потреблять, попадают в кишечник и всасываются в кровоток до того, как они могут быть обезврежены.
Прохождение пищи через тонкую кишку может занять от 30 до 60 минут, так как большая часть пищеварительного тракта движется со скоростью примерно 30 см в минуту. Однако корм обычно проходит через тонкий кишечник за 3-4 часа. Чем быстрее дигеста проходит через тонкий кишечник, тем меньше времени остается на действие ферментов. Было показано, что добавление масла в рацион лошадей уменьшает поток корма через тонкую кишку, что дает пищеварительным ферментам больше времени для переработки крахмалов, белков и жиров, тем самым увеличивая общую усвояемость этих питательных веществ трактом и максимально улучшая пищеварение тонкого кишечника. эффективность. Лошади очень восприимчивы к коликам или смерти от токсичных материалов в корме. В отличие от коровы, у которой в рубце есть бактерии, которые могут обезвреживать материалы до того, как они достигнут тонкой кишки, токсичные материалы, которые лошадь может потреблять, попадают в кишечник и всасываются в кровоток до того, как они могут быть обезврежены. Поэтому очень важно не давать лошадям заплесневелые или испорченные корма. Мочевина представляет собой кормовую добавку, скармливаемую крупному рогатому скоту, которая может быть использована в их рубце для производства белка. Лошади не могут использовать эту кормовую добавку, потому что она всасывается в тонком кишечнике до того, как попадет в слепую кишку, где ее можно было бы использовать. Мочевина может быть токсичной для лошади, но лошадь может переносить уровень, при котором ее добавляют в большинство кормов для крупного рогатого скота. Микробный белок, который синтезируется в толстом кишечнике, не может быть использован лошадью в достаточной степени. Это означает, что животные с высокой потребностью в белке (жеребята, лактирующие кобылы и, возможно, интенсивно тренирующиеся лошади) должны получать высококачественный белок, который может расщепляться и всасываться преимущественно в тонком кишечнике. В практическом смысле это означает не обязательное увеличение содержания сырого протеина в корме для лошадей, а повышение его качества.
Поэтому очень важно не давать лошадям заплесневелые или испорченные корма. Мочевина представляет собой кормовую добавку, скармливаемую крупному рогатому скоту, которая может быть использована в их рубце для производства белка. Лошади не могут использовать эту кормовую добавку, потому что она всасывается в тонком кишечнике до того, как попадет в слепую кишку, где ее можно было бы использовать. Мочевина может быть токсичной для лошади, но лошадь может переносить уровень, при котором ее добавляют в большинство кормов для крупного рогатого скота. Микробный белок, который синтезируется в толстом кишечнике, не может быть использован лошадью в достаточной степени. Это означает, что животные с высокой потребностью в белке (жеребята, лактирующие кобылы и, возможно, интенсивно тренирующиеся лошади) должны получать высококачественный белок, который может расщепляться и всасываться преимущественно в тонком кишечнике. В практическом смысле это означает не обязательное увеличение содержания сырого протеина в корме для лошадей, а повышение его качества. Это может означать, что правильные уровни незаменимых аминокислот, таких как лизин, метионин и треонин, находятся на уровне, достаточном для удовлетворения потребностей лошади.
Это может означать, что правильные уровни незаменимых аминокислот, таких как лизин, метионин и треонин, находятся на уровне, достаточном для удовлетворения потребностей лошади.
Задняя кишка
Задняя кишка или толстая кишка, к которой ее обычно относят, состоит из слепой кишки, толстой (или восходящей ободочной кишки, тонкой кишки, прямой кишки и ануса). длина всего кишечника составляет примерно 7 м, а объем составляет 140-150 л. Пищеварение в задней кишке в основном микробное, а не ферментативное. Пищеварение в задней кишке осуществляется миллиардами симбиотических бактерий, которые эффективно расщепляют растительные волокна и непереваренные крахмалы. на более простые соединения, называемые летучими жирными кислотами (ЛЖК), которые могут всасываться через стенку кишечника По сравнению с пищеварительным трактом жвачных животных лошадь не так хорошо приспособлена к перевариванию продуктов травы с высоким содержанием сырой клетчатки, низким содержанием белка и низким содержанием белка. уровни углеводов, крахмала и жира.Однако они справляются с этим намного лучше, чем человек или свинья!И лошадиные уменьшили эти недостатки, выборочно выпасая большое количество корма каждый день.
уровни углеводов, крахмала и жира.Однако они справляются с этим намного лучше, чем человек или свинья!И лошадиные уменьшили эти недостатки, выборочно выпасая большое количество корма каждый день.
Слепая кишка
Слепая кишка представляет собой слепой мешок длиной примерно 1,2 м, вмещающий около 28–36 литров корма и жидкости. Слепая кишка представляет собой микробный инокуляционный чан, похожий на рубец коровы. Микробы расщепляют корм, который не переварился в тонком кишечнике, особенно волокнистые корма, такие как сено или пастбища. Слепая кишка имеет необычный дизайн, потому что ее вход и выход находятся в верхней части органа. Это означает, что корм поступает сверху, перемешивается и затем выбрасывается вверх. Такая конструкция является причиной проблем, если животное ест много сухих кормов без достаточного количества воды или если происходит быстрая смена рациона. Оба могут вызвать уплотнение в нижнем отделе слепой кишки, что, в свою очередь, вызывает боль (колики). Микробная популяция в слепой кишке несколько специфична в отношении того, какие корма она может переваривать. Микробной популяции слепой кишки может потребоваться до 2-3 недель, чтобы приспособиться к новой диете и вернуться к нормальной функции. Вот почему вы будете читать на этикетках пакетов, чтобы медленно вводить новые корма для лошади в течение 7-14 дней. Корм остается в слепой кишке около семи часов, что дает бактериям время начать его расщепление в процессе ферментации. Микробы будут производить витамин К, витамины группы В, белки и жирные кислоты. Витамины и жирные кислоты будут усваиваться, но белок усваивается незначительно.
Микробная популяция в слепой кишке несколько специфична в отношении того, какие корма она может переваривать. Микробной популяции слепой кишки может потребоваться до 2-3 недель, чтобы приспособиться к новой диете и вернуться к нормальной функции. Вот почему вы будете читать на этикетках пакетов, чтобы медленно вводить новые корма для лошади в течение 7-14 дней. Корм остается в слепой кишке около семи часов, что дает бактериям время начать его расщепление в процессе ферментации. Микробы будут производить витамин К, витамины группы В, белки и жирные кислоты. Витамины и жирные кислоты будут усваиваться, но белок усваивается незначительно.
Большая кишка
Большая толстая кишка состоит из правого и левого вентральных отделов толстой кишки и спинного отдела толстой кишки, имеет длину около 3-3,5 м и вмещает 86 литров. Микробное пищеварение (ферментация) продолжается, и большая часть питательных веществ, полученных в результате микробного пищеварения, поглощается здесь, а также витамины группы В, вырабатываемые бактериями, и некоторые микроэлементы и фосфор. Брюшная кишка имеет «саккулированную» структуру, напоминающую ряд мешочков. Такая конструкция облегчает переваривание большого количества волокнистых материалов, но из-за своей конструкции может стать большим фактором риска возникновения колик. Пакеты могут легко скручиваться и наполняться газом из-за ферментации корма. Корм может добраться сюда всего за семь часов и останется здесь на 48-65 часов.
Брюшная кишка имеет «саккулированную» структуру, напоминающую ряд мешочков. Такая конструкция облегчает переваривание большого количества волокнистых материалов, но из-за своей конструкции может стать большим фактором риска возникновения колик. Пакеты могут легко скручиваться и наполняться газом из-за ферментации корма. Корм может добраться сюда всего за семь часов и останется здесь на 48-65 часов.
Тонкая кишка, прямая кишка и задний проход
Малая кишка примерно такой же длины, как и толстая кишка, но имеет диаметр всего около 10 см. К настоящему времени подавляющее большинство питательных веществ уже переварено, а то, что осталось, не может быть переварено или использовано лошадью. Основная функция тонкой кишки заключается в утилизации избыточной влаги и возвращении ее в организм. В результате образуются фекальные шарики. Эти фекальные шарики, которые представляют собой непереваренную и в основном неперевариваемую часть того, что было съедено 36-72 часа назад, затем попадают в прямую кишку и выводятся в виде навоза через задний проход.
Пищеварительный канат
Желудочно-кишечный тракт лошадей хорошо функционирует при нормальных постоянных условиях. Однако, как известно всем, кто занимается лошадьми, желудочно-кишечный тракт лошадей чрезвычайно чувствителен и легко нарушается, а колики являются причиной номер один гибели лошадей. Любое резкое изменение в рационе может поставить под угрозу и изменить популяцию бактерий в задней кишке лошади, что может привести к коликам и, по крайней мере, снижению эффективности пищеварения диеты. Поддерживать микрофлору в хорошем состоянии может быть сложно, если лошадь находится в состоянии стресса, путешествует на большие расстояния, перенесла болезнь или травму, получила антибиотики, отлучила жеребенка или высокопроизводительную лошадь кормят большим количеством зерна. Крайне важно, чтобы мы относились к задней кишке лошади с уважением и следили за рационом наших лошадей и их общим состоянием здоровья. Пытаясь кормить своих лошадей как можно ближе к их естественному пастбищному привычку (чаще небольшими порциями), вы значительно снизите риск заболеваний желудочно-кишечного тракта.
Leave A Comment